Leaf consumption defines success of the protocol. Healthy, accurately staged larvae should begin feeding immediately after placement on the leaf surface and feeding should continue in a fairly consistent manner throughout the infestation time. In Video 1, the larva at the top begins to chew immediately after placement and maintains a consistent rate while feeding. This is especially important if assaying early gene expression events after infestation. The larva at the bottom did not consume any leaf material and is an example of an unsuccessful infestation.
Visual approximations during leaf consumption are monitored to ensure enough damage is produced. The amount of leaf material consumed can be calculated as the percentage of damage by using images of leaves before and after the infestation. Figure 2 illustrates the variable rates of consumption by different developmental stages of M. sexta larvae on different types of potato plants. The leaves in Figure 2A were detached from 2-week-old nodal-propagated tissue culture plants. All the leaves in this figure are of the same size but were subjected to infestation by different stages of larvae for 5 min. Figure 2A: I-IV, Video 2, Video 3, Video 4, and Video 5 illustrate the different rates of consumption and feeding styles for each larval stage from a portion of the infestation. This is helpful in determining how much damage is possible using each leaf and larval stage combination and illustrates the voracious nature of the older larvae. The use of 1st instar larvae is not recommended as their mandibles are not developed enough to produce damage in the 5 min infestation window. It is important to note that younger more tender leaves derived from tissue-culture-grown plants are often more palatable to larvae. Based on these results, 4th instar larvae were chosen to further assess damage to leaves from more mature, soil grown potato plants. The soil grown plants more closely approximate those acquired in the field. Figure 2B illustrates the range of damage when 3rd and 4th instar larvae were applied to leaves detached from soil-grown potato plants. Two-week-old nodal propagated plants were transplanted to soil and grown for an additional 3 weeks. Damaged leaves from soil grown plants fell into three groups; 15-20%, 20-30%, and 30-40%. The three leaves in each group are shown to represent the different levels of damage for each range. The leaves from more mature soil-grown plants needed more larvae applied and a longer infestation time to reach the same level of damage observed in the leaves from younger nodal-propagated plants. These results illustrate the range of outcomes possible from successful herbivory from different types of plants.
After successful herbivory is complete and percent damage is assessed, leaf tissue is assayed for gene expression. Gene expression results should indicate a robust induction or repression of transcripts involved in the response to infestation. Figure 3 illustrates the gene expression profiles of six C2H2 transcription factors from a successful herbivory experiment in detached leaves. C2H2 zinc finger StZFP2 was induced by M. sexta herbivory in potato in previous whole plant studies24. Although StZFP2 was induced by herbivory in the detached leaf assay, infestation was not significant when compared to the effect of the detachment itself. However, two other StZFP2 homologs, StZFP5 and StZFP7, were significantly induced at 40 and 80 min post herbivory when compared to detached controls. StLOX3 and StMYC2 are marker genes induced by both wounding and herbivory and indicate the involvement of jasmonic acid regulated defense pathways. The goal of this particular study was to identify infestation-responsive transcription factors among a panel of candidate genes. The identification of two C2H2 zinc finger transcription factors that are robustly responsive to M. sexta herbivory supports the use of detached leaves for infestation assays.

Figure 1: Flowchart for the infestation protocol. Schematic representation of the steps included in the experimental workflow of the infestation section of the protocol. Please click here to view a larger version of this figure.

Figure 2: Leaf damage caused by M. sexta larvae. (A) Percentage damage to tissue culture leaves at each larval stage over 5 min. Roman numerals I-IV refer to videos 2-5 respectively that show each instar feeding on the leaf pictured. (B) Range of damage to soil-grown leaves by 4th instar over 20 min. Please click here to view a larger version of this figure.
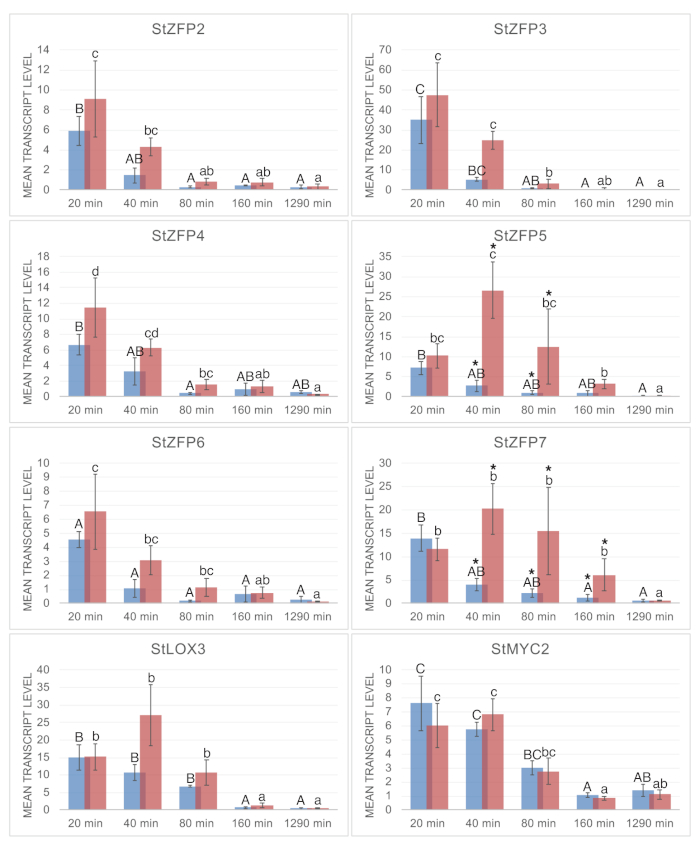
Figure 3: Gene expression analysis of leaves. Real-time quantitative polymerase chain reaction (RT-qPCR) gene expression analysis of C2H2 zinc finger transcription factors is shown. Mean transcript level is, 2−ΔCT with (ΔCT = CT of test gene – CT of exogenous control gene). Excised control leaves in (blue) and excised infested leaves in (red) are shown at each time point. Each value is the average of three biological replicates. Three-way ANOVA was conducted on the ΔCt values. Significant differences in control leaves over time are indicated with capital letters. Significant differences in infested leaves over time are indicated in lower case letters. Significant differences between control and infested leaves at the same time point is shown with an asterisk (*). Pr > F values were all less than 0.001, except for StZFP6 control treatment with 0.0086. Error bars represent standard deviation. NCBI accession numbers are: StLOX3-X96406.1, StZFP2- MK809525, StZFP4-CV500970.1, StZFP6-DN587601.1, StZFP7-DN590005.1. Spud DB accession numbers from <http://solanaceae.plantbiology.msu.edu> are StMYC2-PGSC0003DMT400045204, StZFP3-PGSC0003DMT400040144, StZFP5-PGSC0003DMT400040141. This figure has been modified from22. Please click here to view a larger version of this figure.

Video 1: Video clip of larval feeding. Second instar M. sexta larvae feeding (top) and not feeding (bottom) on a leaf from two week old tissue culture grown potato plant. Please click here to view this video. (Right-click to download.)

Video 2: Figure 2A video clip (I) of larval feeding. Second instar M. sexta larva feeding on a leaf from two week old tissue culture grown potato plant. Please click here to view this video. (Right-click to download.)

Video 3: Figure 2A video clip (II) of larval feeding. Third instar M. sexta larva feeding on a leaf from two week old tissue culture grown potato plant. Please click here to view this video. (Right-click to download.)

Video 4: Figure 2A video clip (III) of larval feeding. Fourth instar M. sexta larva feeding on a leaf from two week old tissue culture grown potato plant. Please click here to view this video. (Right-click to download.)

Video 5: Figure 2A video clip (IV) of larval feeding. Fifth instar M. sexta larva feeding on a leaf from two week old tissue culture grown potato plant. Please click here to view this video. (Right-click to download.)
