After one week (7-10 days) in the chip with differentiation media, NSCs differentiate into neurons and neuronal projections enter the axonal compartment (Figure 3). Within the chip, neurons attach and distribute evenly within the somatic compartment. In comparison, neurons in PDMS devices clump/aggregate as early as 5 days post addition of differentiation media, leading to compromised cell health as can be seen in the Figure 3B (day 13 magnified image). Neurons in the chips look healthy with bundled axons. Healthy neurons can be maintained within the chips for 4-5 weeks.
To visualize neuron maturation and dendritic spine development, modified rabies virus was delivered to the axonal compartment at Day 29 to retrograde label neurons, including dendritic spines, with mCherry fluorescent protein. Four days after rabies virus infection, neurons extending axons into the axonal compartment expressed mCherry. The neurons at differentiation day 33 showed formation of dendritic spines (Figure 4). Visualization of dendritic spines demonstrates that NSC derived neurons differentiated within the chips form mature synapses.
The multi-compartment chip is also compatible with immunocytochemistry to visualize the cellular localization of protein. After 26 days of maintaining neurons in differentiation media, neurons were labeled for the excitatory synaptic marker, vGlut1 (Figure 5). These results show that virally labeled neurons co-localize with vGlut1 (Figure 5E) and neuron specific marker, β-tubulin III (Figure 5F-H).
The ability to create distinct microenvironments isolated to axons was also demonstrated using Alexa Fluor 488 hydrazide, a low molecular weight fluorescent dye (Figure 6).
Axon injury studies are commonly performed within compartmentalized devices. A proof of principle experiment was performed to selectively injure axons of differentiated neurons with the pre-assembled COC chip (Figure 7). Results were equivalent to silicone compartmentalized devices6,13,14.

Figure 1: The pre-assembled COC, multicompartment microfluidic chip. (A) A drawing of the chip identifying the upper and lower wells. The size of the chip is 75 mm x 25 mm, the size of standard microscope slides. (B) A zoomed in region showing the channels and microgrooves separating the channels. Additional details are provided in Nagendran et al.12. (C) This photograph illustrates the creation of isolated microenvironments within each compartment using food coloring dye. This entire figure has been reproduced from 12. Please click here to view a larger version of this figure.
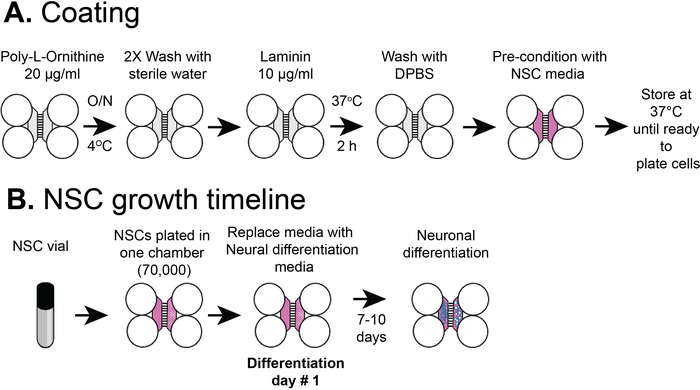
Figure 2: Plastic compartmentalized chip coating and NSC cell plating timeline. (A) Plastic multicompartment chips were coated with a pre-coating solution, and then poly-L-ornithine and laminin before pre-conditioning with NSC media. (B) NSCs were plated with 7 x 104 cells in the somatic compartment of the chip. The cells were grown in NSC media for 24 h and then the media was replaced with neural differentiation media. Differentiated neurons were observed by 7-10 days post differentiation. Please click here to view a larger version of this figure.

Figure 3: hSC-neuron growth comparison in chips and PDMS devices. (A) A phase contrast image of hSC-neurons grown at 13 days after differentiation in the plastic multicompartment chip. (B) A zoomed in region of hSC-neurons cultured within the white box in (A) and an equivalent region within a PDMS device (right). hSC-neurons within the chips attach well. Aggregated neuron clusters form in PDMS-based devices. Representative of 2 independent experiments. Please click here to view a larger version of this figure.
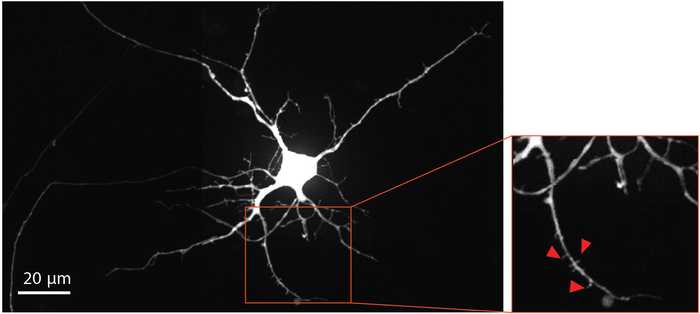
Figure 4: Human NSC derived neurons show dendritic spine morphology. Retrograde labeled mCherry neuron at differentiation day 33 grown within the chip. The magnified region outlined in red, highlights the presence of dendritic spines, which provide evidence supporting the development of mature glutamatergic synapses. Red arrows point towards dendritic spines. Please click here to view a larger version of this figure.
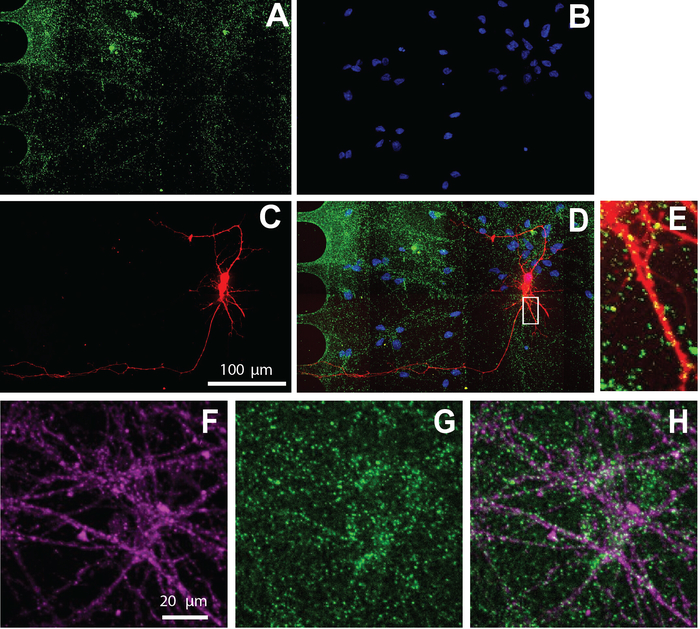
Figure 5: Human NSC derived neurons cultured within chips exhibit excitatory synapses. Immunostaining was performed at differentiation day 26. Neuron imaging was performed in the somatic compartment. A) vGlut1 (green) and (B) DAPI (blue) immunolabeling. (C) mCherry-labeled neurons (red) retrograde labeled using a modified rabies virus. (D) A merged fluorescent micrograph of (A-C). (E) Dendritic spines and vGlut1 positive puncta colocalized with mCherry-positive dendrites, shown in a zoomed in region from (D). Immunofluorescence micrographs of (F) neuron specific marker, β-tubulin III (magenta), and (G) vGlut1 (green). (H) Overlay of β-tubulin III, and vGlut1. Please click here to view a larger version of this figure.

Figure 6: Virally transduced mCherry neurons extend projections into an axon localized microenvironment established within the preassembled COC chip. (A) mCherry labeled neuron extend axons through the microgrooves of the chip and into an isolated axon compartment. Isolation of the axon compartment is visualized using Alexa Fluor 488 hydrazide. Imaging of neurons occurred at differentiation day 26 and 3 days after infection with the modified rabies virus. (B) The fluorescence image in (A) is merged with a DIC image. Note the location of the microgrooves. Please click here to view a larger version of this figure.
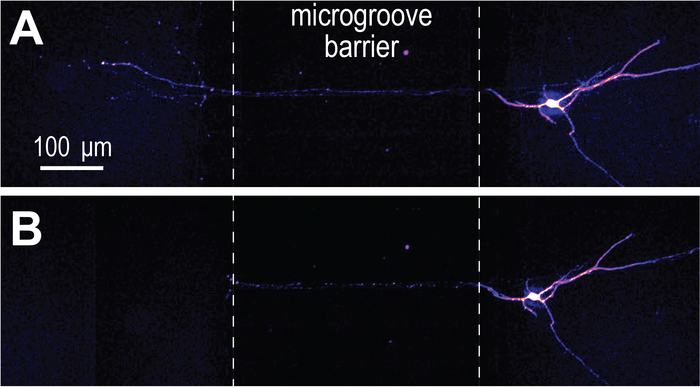
Figure 7: Axotomy performed within the COC multicompartment chip. (A) mCherry labeled neurons at differentiation day 33 were imaged before axotomy. ‘Fire’ color look-up table. (B) Immediately after axotomy, showing that axons are completely severed. Please click here to view a larger version of this figure.
| Plastic multi-compartment chips | PDMS multi-compartment devices |
| isolate axons | isolate axons |
| establish microenvironments | establish microenvironments |
| axotomize neurons | axotomize neurons |
| optically transparent | optically transparent |
| compatible with high resolution imaging | compatible with high resolution imaging |
| compatible with fluorescence microscopy | compatible with fluorescence microscopy |
| fully assembled | assembly to substrate required |
| healthy axons >21 days | healthy axons >14 days |
| hydrophilic culturing surface | hydrophobic |
| gas impermeable | gas permeable |
| rounded microgrooves and channels | straight microgrooves |
| not compatible with laser ablation | Can be used for laser ablation when PDMS chambers are assembled on special laser ablation compatible slides. |
| device cannot be altered to remove top | top is removable for staining within microgrooves |
| not compatible with mineral oil-based immersion oils (silicone-based oils are fine) | compatible with mineral oil-based immersion oils |
| impermeable to small molecules and organic solvents | absorption of small molecules & organic solvents |
| No leakage issues with the chip | coating with poly-L-ornithine and laminin make the devices leaky |
| differentiated neurons remain evenly distributed (> 4 weeks) at the tested cell densities | differentiated neurons begin to aggregate after 3-4 days in culture at the tested cell densities |
Table 1: Comparision of multi-compartment COC chips and silicone devices for culturing neurons
