This section provides representative results for ERG measurements taken daily from 4 to 7 dpf. From 4 dpf, ERG responses show robust a- and b-wave components, which arise from photoreceptors and bipolar cells, respectively. At each age tested, the amplitude of the b-wave increased with light intensity (Figure 2; Figure 3). Notably, the sensitivity of the larval zebrafish retina to dimmer flashes increased with age. The a- and b-wave were not recognizable at intensities lower than -1.61 log cd.s/m2 at 4 dpf, whereas clear signals were detectable at these intensities for older larvae (Figure 2). The b-wave response grew substantially between 4 and 5 dpf (P < 0.0001; Figure 2A-B; Figure 3B). Although the b-wave at lower intensities showed little change between 5 and 7 dpf, the signal at 2.48 log cd.s/m2 was greater at 7 dpf compared with 5 and 6 dpf (P < 0.0001; Figure 2; Figure 3B). A- and b-wave implicit times became significantly faster after 5 dpf (P < 0.0001; Figure 3C-D). Overall, these results demonstrate maturation of zebrafish retinal function between 4 to 7 dpf. Interestingly, the a-wave amplitude appeared to decrease from 5 to 7 dpf (Figure 3A). This may be because the maturation of synaptic connections in the outer retina shortens the latency of bipolar cells responses, resulting in faster b-wave onset that masks the a-wave. Those wishing to study the a-wave can employ pharmacological treatment to block post-photoreceptoral responses (i.e. the b-wave component).
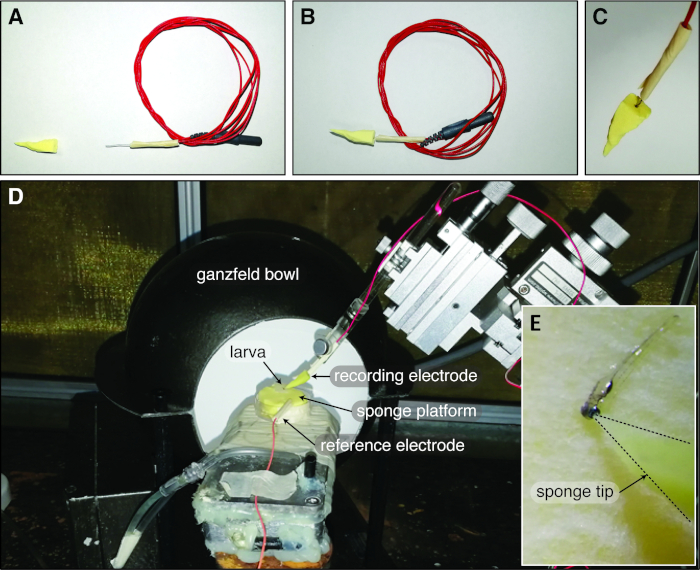
Figure 1: Zebrafish Ganzfeld ERG set up with the cone-shaped sponge-tip electrode. (A) The cone-shaped sponge tip and the chlorinated silver electrode are air dried before constructing the sponge-tip electrode. (B-C) Subsequently, the chlorinated silver wire is inserted into the sponge cone through the base to form the complete electrode. (D) In the typical larval zebrafish Ganzfeld ERG setup, the reference electrode is inserted into the sponge platform and the zebrafish larva is covered by the Ganzfeld bowl. (E) The sponge-tip electrode gently touches the central corneal surface of the larval eye. Please click here to view a larger version of this figure.
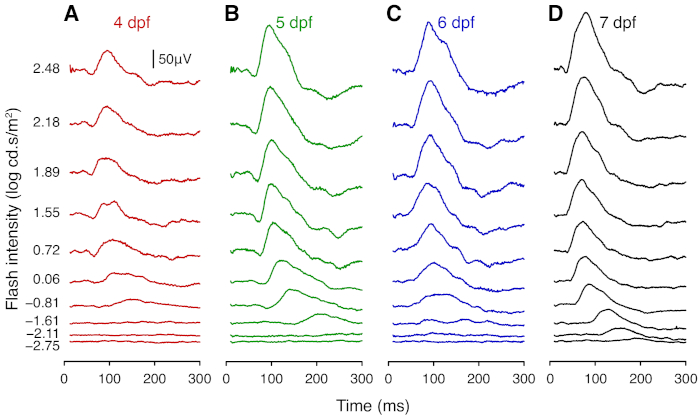
Figure 2: Representative average ERG traces of wild-type larval zebrafish. Average ERG traces of wildtype zebrafish at (A) 4 dpf (n = 8), (B) 5 dpf (n = 8), (C) 6 dpf (n = 7), and (D) 7 dpf (n = 9). Responses were elicited using flashes from white LEDs. At each age, the traces show responses to flashes of (from bottom to top) -2.75, -2.11, -1.61, -0.81, 0.06, 0.72, 1.55, 1.89, 2.18, 2.48 log cd.s/m2. Scale bar = 50 µV. Please click here to view a larger version of this figure.
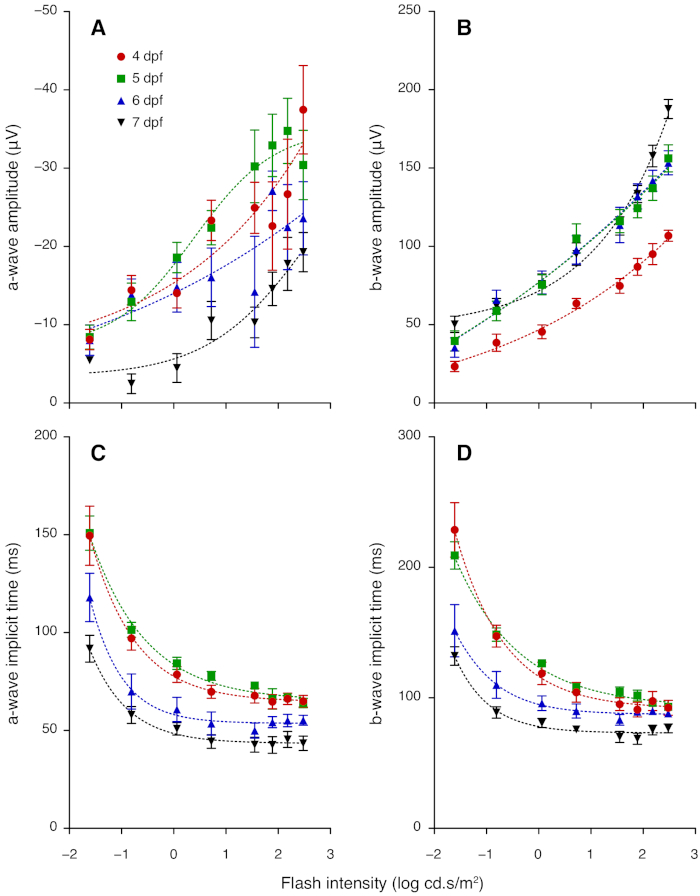
Figure 3: ERG a- and b-wave amplitudes and implicit times for 4 to 7 dpf zebrafish. (A) Group average (± standard error of the mean) a-wave amplitude increased with flash intensity but decreased with age in 4–7 dpf larvae. (B) Average b-wave amplitude in 4–7 dpf larvae increased with flash intensity; amplitude grew between 4 and 5 dpf. (C) Average a-wave implicit time and (D) average b-wave implicit time became faster between 5, 6 and 7 dpf. Lines of best fit are derived from non-linear regression. Please click here to view a larger version of this figure.
| Stimulus light intensity (log cd.s.m-2) | Number of repeats | Inter-stimulus interval (s) |
| -2.75 | 3 to 6 | 10 |
| (30 s before next) | ||
| -2.11 | 3 to 6 | 10 |
| (30 s before next) | ||
| -1.61 | 3 to 6 | 10 |
| (30 s before next) | ||
| -0.81 | 3 to 6 | 10 |
| (60 s before next) | ||
| 0.06 | 3 to 6 | 10 |
| (60 s before next) | ||
| 0.72 | 1 to 3 | 60 |
| 1.55 | 1 to 3 | 60 |
| 1.89 | 1 to 3 | 60 |
| 2.18 | 1 to 3 | 60 |
| 2.48 | 1 to 3 | 60 |
Table 1: Example protocol of ERG recordings. Stimulus presentations start from the dimmest (top) and progress to brighter (bottom) light levels, with progressively longer inter-stimulus intervals to ensure that dark adaption is maintained. The number of signals averaged at each intensity depends on the signal-to-noise level.
