Using the procedure described, primary human monocytes from HIV-infected individuals and healthy donors were isolated. All data presented here were obtained from HIV+ subjects undergoing cART with low (<20 copies/mL) or undetectable viral loads and normal CD4+ T cell counts. Immediately after isolation, cells were stained, and flow cytometry was performed to confirm the purity of cell populations. Results showed that >97% of cells stained positive for CD14 (data not shown). For polarization of macrophages, a published protocol was used28. Primary human monocytes were cultured in the presence of GM-CSF or M-CSF for 6 days. On the sixth day, cells were activated towards either M1 or M2 macrophages. Twenty-four hours post-activation, cells were harvested and stained for flow cytometry analysis of macrophage cell markers.
Figure 1 shows representative histograms of M1-activated control- (Figure 1A) and patient-derived (Figure 1B) cells with increased levels of CD80 and CD83 and decreased levels of CD163 when compared to non-activated T0 cells, as well as M2-activated cells with increased levels of CD163 and CD209. Panel C in shows expression of CD80, CD83, CD163, and CD209 in M1 and M2 polarized cells. The graph represents the average data obtained from three control- and three HIV-derived sets of cells. As expected, expression levels of CD80 and CD83 increased in M1 compared to M2 polarized cells, while CD209 and CD163 were more highly expressed in M2 compared to M1 polarized cells. Interestingly, CD80 and CD83 appeared to be more highly expressed in control-derived cells compared to HIV-derived cells. However, potential differences in the ability to polarize and/or expression levels of polarization markers in HIV-derived cells compared to controls requires further investigation. Although GM-CSF, M-CSF, LPS, and IFN-γ were chosen as treatments, other combinations of growth factors, cytokines, or stimulators may be used with this protocol28.
After preliminary experiments showed that the isolation procedure was successful, freshly collected monocytes were plated in 6 well plates and transfected with a scrambled, near-infrared-labeled miRNA to determine transfection efficiency. Cells were imaged 24 h post-transfection by fluorescent confocal microscopy. With this method, >90% efficiency of transfection was achieved, as determined by flow cytometry (Figure 2A) and confocal microscopy (Figure 2B).
Next, viability of the cells after transfection was determined. Figure 3 shows the viability of cells, determined using a colorimetric assay as an average of cells derived from two patients (Figure 3A) and two controls (Figure 3B) transfected at day 1 (Figure 3A) or day 4 (Figure 3B) and harvested at day 7. For this experiment, 50,000 cells were plated on a 96 multi-well plate and transfected following the protocol. In general, transfection did not significantly reduce the viability of cells, regardless of the stage of maturation of the cells and the transfection conditions (i.e., mock, scrambled siRNA/miRNA, or siRNA/miRNA). It should be noted that Figure 3 represents data obtained from patient-derived cells (panel A) and control-derived cells (panel B). This was necessary, since not enough cells to perform the full experiment (both viability and western blot for day 1 and day 4 transfections, in six different conditions per transfection) with a single sample were able to be obtained. Nevertheless, the figure provides representative results obtainable with either control- or patient-derived cells.
Then, the effectiveness of siRNA transfection on target mRNA was assessed by evaluating protein expression. Results in Figure 4 show effective downregulation of EIF4EBP1, a translational regulator highly abundant in these cells, upon transfection with a specific siRNA at both day 1 (Figure 4A) and day 4 (Figure 4C). The same regulator also maintained expression under the various control conditions (i.e., untransfected, mock, scrambled siRNA, and EIF4EBP1 siRNA without transfection reagent: siR*). Quantification of western blot experiments for day 1 and day 4 transfection are shown in Figure 4B and Figure 4D, respectively. Additionally, expression levels of miR-146a-5p following transfection at day 1 or day 4 by RT-qPCR of HIV-derived cells were determined (Figure 4E). Cells transfected with miRNA mimic showed a 48- to 72-fold increase in miRNA expression over untransfected cells, while all transfection controls show no appreciable changes.
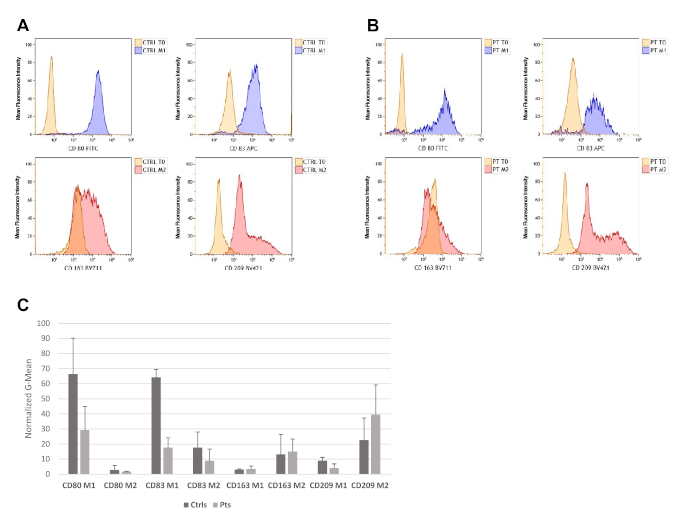
Figure 1: Monocyte-derived macrophages are successfully polarized and activated towards M1 or M2 macrophage phenotypes. Flow cytometry analysis results of cells derived from one healthy control (A) and one HIV-derived cell sample (B) show levels of CD80, CD83, CD163, and CD209. The experiment was repeated with two additional controls and two additional HIV-positive samples with similar results. Cell population of interest was gated on the basis of forward and side scatter parameters, followed by doublet discrimination. (C) Bar graph showing CD80, CD83, CD163, and CD209 in M1 and M2 polarized cells. The graph represents the average data and standard deviations obtained from three control- (Ctrl) and three HIV-derived (Pts) sets of cells. Please click here to view a larger version of this figure.
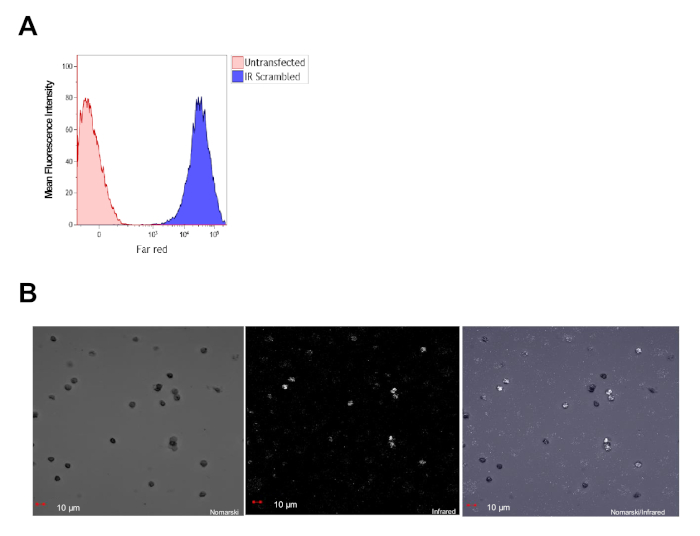
Figure 2: Primary human CD14+ cells are efficiently transfected with miRNAs. (A) Flow cytometry analysis of primary monocytes derived from healthy controls, 24 h post-transfection, showing >90% transfection efficiency. (B) Representative confocal image taken 24 h post-transfection shows that all cells in the field express the miRNA conjugated with a near-infrared dye (in white). Please click here to view a larger version of this figure.
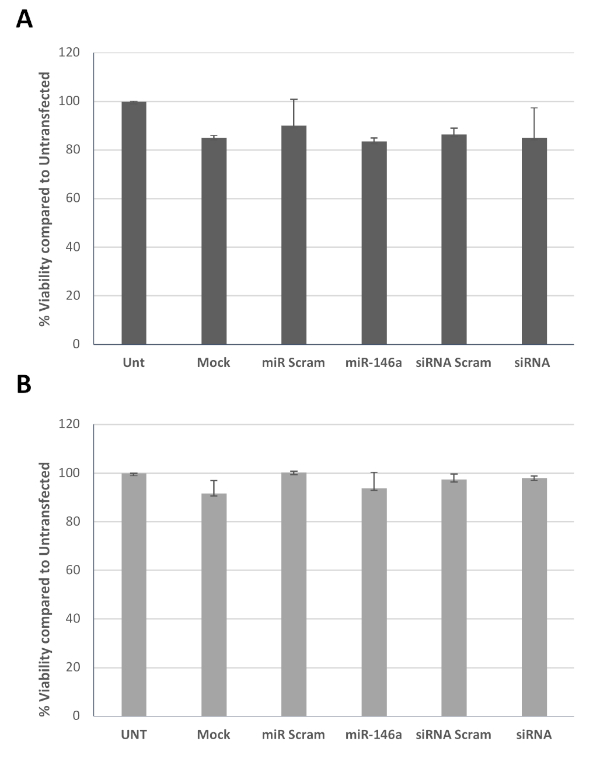
Figure 3: Viability of transfected cells. Graph bars represent average cell viability of two HIV+ patients (A) and two healthy controls (B) determined using a colorimetric assay, after transfection with miR-146a-5p or siRNA to EIF4EBP1 (siRNA) and the appropriate controls at day 1 (A) or day 4 (B), all tested at day 7. Please click here to view a larger version of this figure.
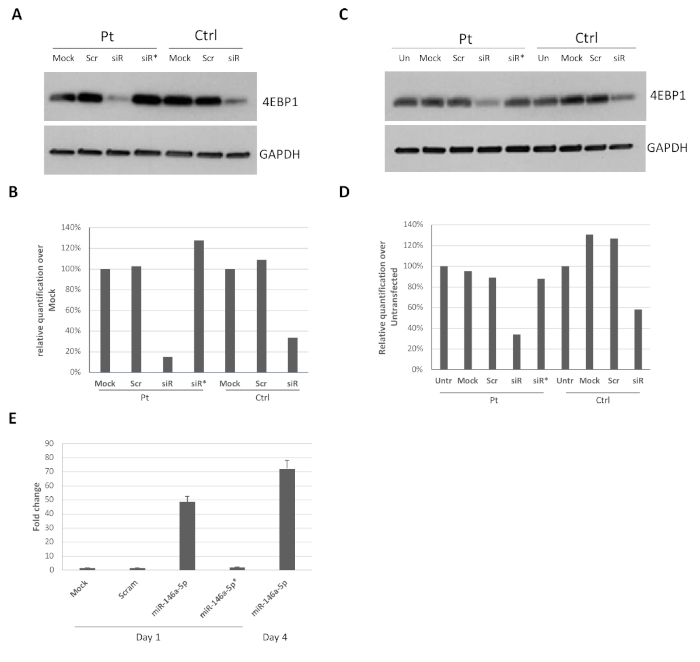
Figure 4: Efficient downregulation of protein levels upon siRNA/miRNA transfection post-isolation of CD14+ monocytes. Control and HIV-derived monocytes (1 x 106 cells) were transfected at day 1 (A) or day 4 (C) post-isolation using siRNA against EIF4EBP1 mRNA. Panels (B) and (D) represent quantification of EIF4EBP1 expression compared to GAPDH and is expressed as the percentage over mock for patient (Pt) or control (Ctrl) (A) or untransfected for patient or control (B). (E) Representative bar graph of two experiments showing miR-146a-5p expression in HIV-derived cells transfected at day 4 (bars 1-4) or day 1 (right bar) and harvested at day 7. The fold change is calculated over the untransfected sample (siR = siRNA). siR* or miR-145a-5p* indicate incubation of the cells with siRNA or miR-146a-5p without the transfection reagent. The experiment was repeated 2x with control-derived cells and produced essentially the same results (data not shown). Please click here to view a larger version of this figure.
