When a fluorescence vascular marker is perfused in vascular lumen (here WGA conjugated with Alexa Fluor-488), it is possible to visualize whole vascular trees as shown in Figure 5 (Left panel) using high-speed confocal microscope. Further magnification enables the imaging of capillary in detail (Figure 5, Right Panel). Since the pressurized system supports a constant monitoring of luminal pressure, this preparation can be used for associate changes in arterial diameter with arterial pressure. Video 1 shows that when pinacidil, an ATP-sensitive K+ channel (KATP) agonist was served from lumen, the diameter of the arterioles was increased. Figure 6 shows that when vasoconstrictor ET-1 was applied from the lumen, the diameter of the arteriole was decreased, and the luminal pressure was increased when the flow was set constant.
Due to high-resolution capabilities of confocal microscopy in combination with specific cell markers, this procedure can also be used to visualize many other cells types associated with the microcirculation. Here, we used a mouse (NG2DsRedBAC transgenic mouse) that expresses DsRed fluorescence protein under a pericyte specific promoter (NG2) and labeled the vessels with WGA-Alexa Fluor 488. This allows us to image simultaneously both the capillary (green) and pericytes (red) in the mouse papillary muscle (Figure 7) under conditions that better mimic physiology in live animals.
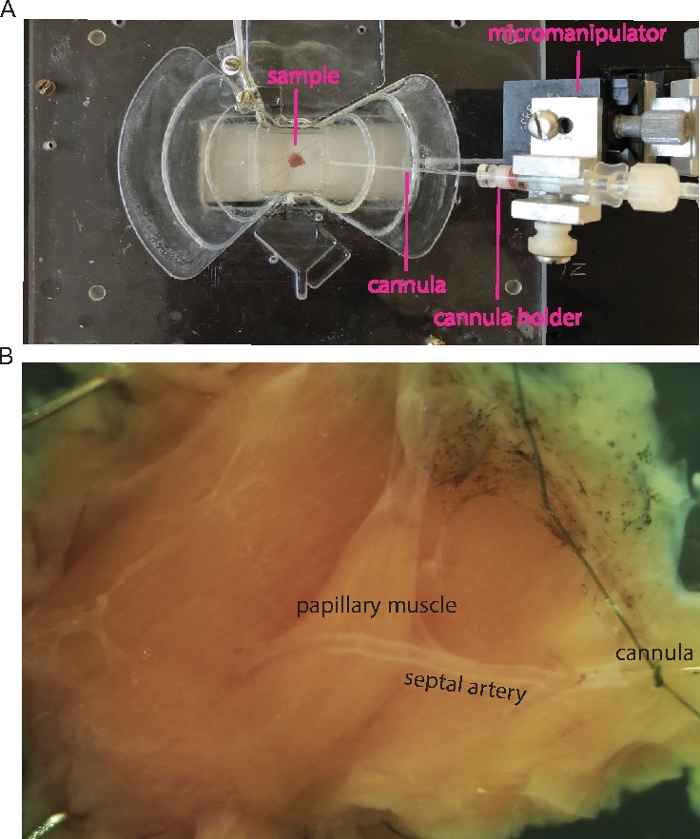
Figure 1: Cannulation of the mouse septal artery.
(A) Transmitted light image shows an example of the cannulated papillary muscle. Micromanipulator, cannula and sample (septum with right ventricle papillary muscle) are indicated as labels. (B) Zoomed-in sample in A shows the papillary muscle and the cannulated septal artery. Please click here to view a larger version of this figure.
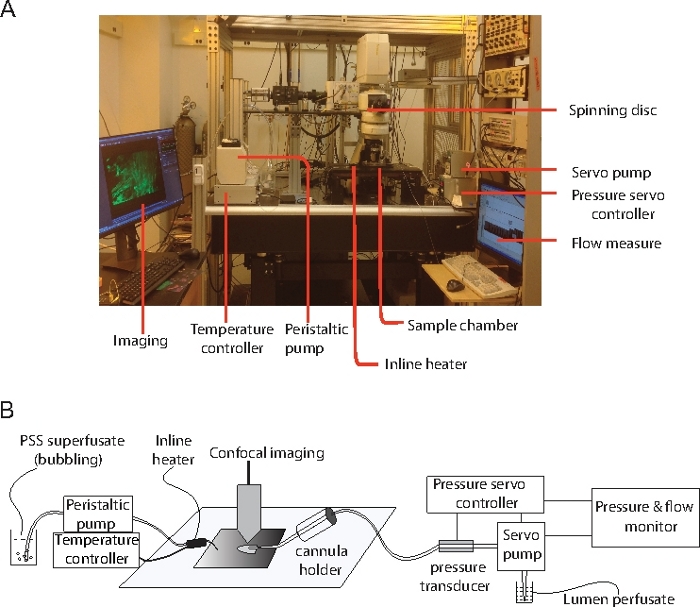
Figure 2: Equipment that was used in all the experiment.
(A) The main components of the setup that are used in the experiment. (B) Diagram illustrates the connections between the papillary muscle preparation and the physiological control experimental equipment. Please click here to view a larger version of this figure.
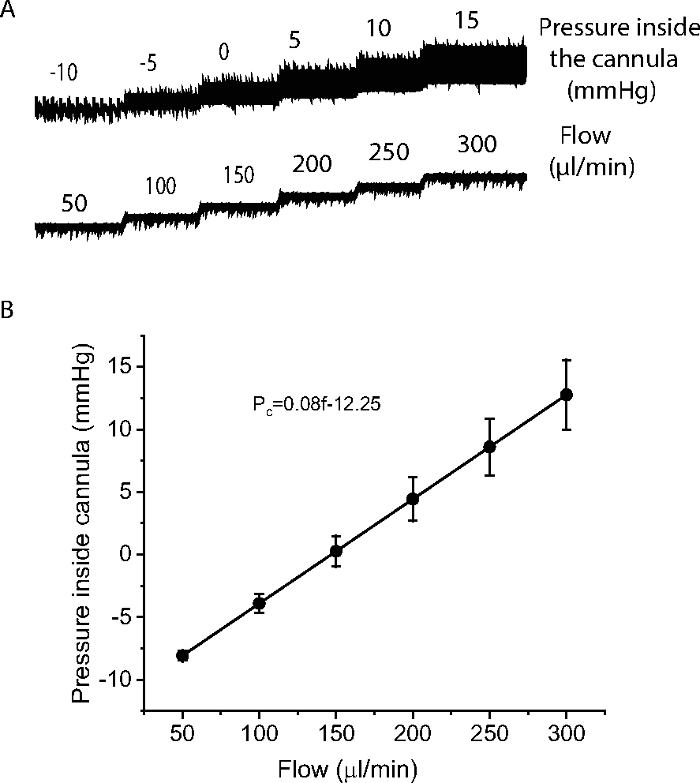
Figure 3: Measurement of pressure inside the cannula.
(A) An example to show how a cannula resistance was determined by measuring pressure inside the cannula over a range of flow (50-300 µl/min). (B) Relationship of the pressure inside the cannula with flow from 6 cannulae. The pressure inside the cannula was proportional to the flow and was fit by the expression Pc=0.08f-12.25, where Pc as pressure inside the cannula, f as flow. N=6 cannulae. Please click here to view a larger version of this figure.
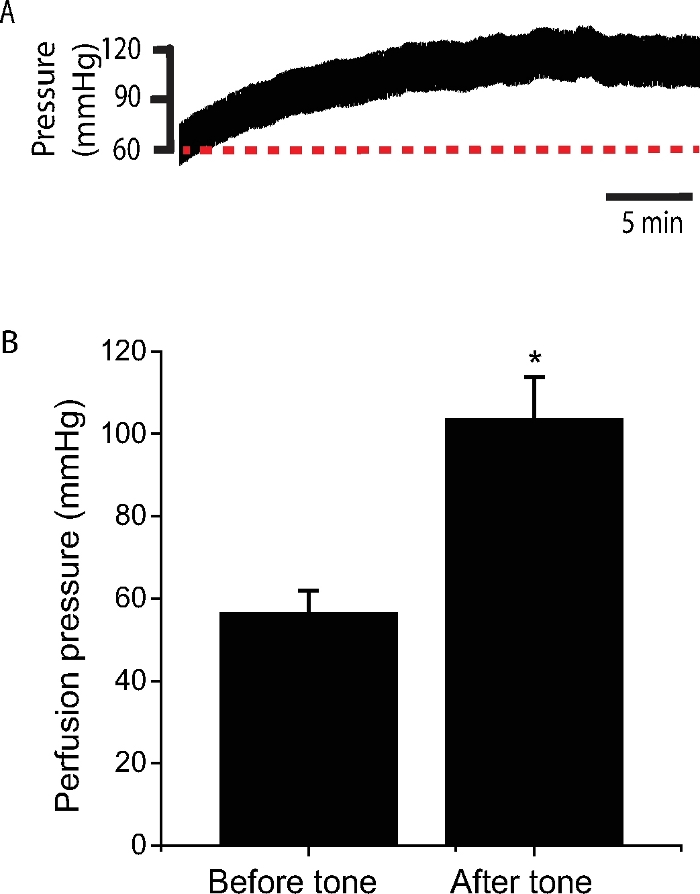
Figure 4: Perfusion pressure change during stabilization at a constant flow.
(A) A typical recording of perfusion pressure during stabilization at a constant flow (~250 µL/min). Note the increase of perfusion pressure after 30 min of stabilization. (B) Statistics show the perfusion pressure before and after the stabilization when tone was developed. The average flow of the arterioles to maintain the initial pressure (~60 mmHg) is 201.7 ± 8.6 µL/min (n= 45 mice). Please click here to view a larger version of this figure.
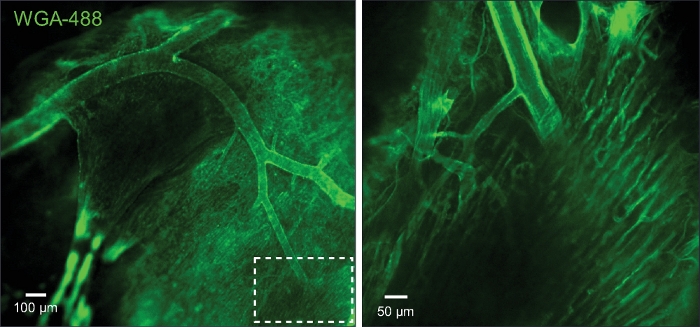
Figure 5: Imaging of capillaries and arterioles.
(A) The image shows the arterioles and capillaries that were loaded with wheat germ agglutinin (WGA). (B) Zoom in of the boxed area in A. Please click here to view a larger version of this figure.
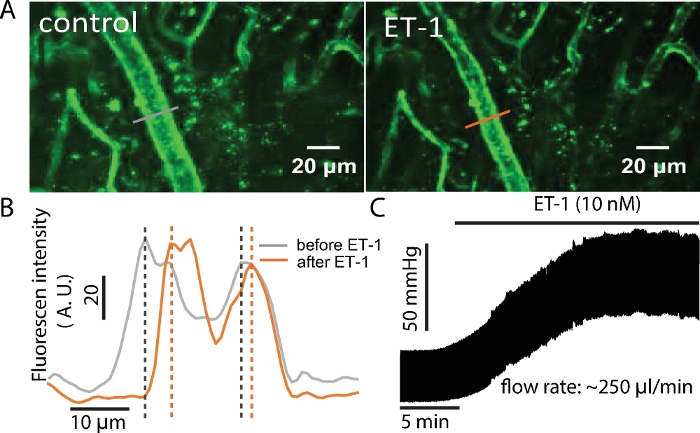
Figure 6: ET-1 increases luminal pressure and decreases arterial diameter.
(A) The arterial diameter changed with the application of ET-1 (10 nM). (B) The WGA fluorescence profiles showing the diameter change by ET-1. The diameter of the arteriole was reflected by the distance between the peak intensity of the fluorescence on the arteriole wall. (C) The luminal pressure increased in the presence of ET-1 (10 nM) at a constant flow. Please click here to view a larger version of this figure.
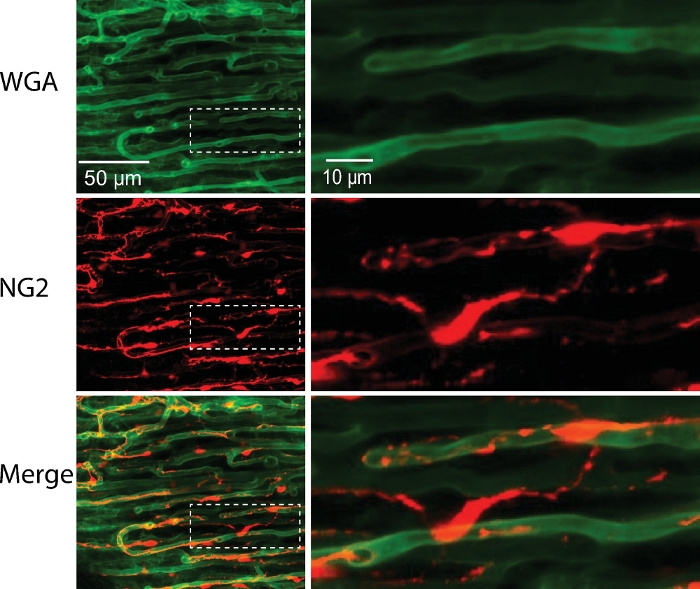
Figure 7: Capillary with pericytes from NG2-DsRed mouse.
Cardiac pericytes (red) and capillaries (green) were imaged in pressurized (40 mmHg) and perfused mouse right ventricular papillary muscle. Right panel, the zoomed-in images of the boxed areas in the left panels. Please click here to view a larger version of this figure.
Video 1: Pinacidil-induced vasodilation. Pinacidil (100 µM) was applied from the lumen. Vasodilation was seen in the arterial tree. Please click here to download this video.
| The composition of physiological saline solution (PSS) | |||
| Reagents | Final concentration (mM) | Molecular weight | g/10 Liters |
| NaCl | 112 | 58.44 | 65.45 |
| KCl | 5 | 74.55 | 3.73 |
| MgSO4 | 1.2 | 120.37 | 1.44 |
| NaH2PO4 | 1.2 | 119.98 | 1.44 |
| NaHCO3 | 24 | 84.01 | 20.16 |
| glucose | 10 | 180.16 | add 1.8 g glucose to 1 Liter PSS before use |
| CaCl2 | 1.8 | 110.99 | add 1.8 ml 1 M CaCl2 to 1 Liter PSS before use |
| The composition of Tyrode's solution | |||
| Reagents | Final concentration (mM) | Molecular weight | g/L |
| NaCl | 140 | 58.44 | 8.18 |
| KCl | 5 | 74.55 | 0.37 |
| NaH2PO4 | 0.33 | 119.98 | 0.04 |
| HEPES | 10 | 238.3 | 2.38 |
| glucose | 5.5 | 180.16 | 0.99 |
| CaCl2 | 1.8 | 110.99 | add 1.8 ml 1 M CaCl2 solution |
| MgCl2.6H2O | 0.5 | 203.3 | add 0.5 ml 1 M MgCl2 solution |
| Note: Adjust pH to 7.4 with 1 M NaOH. | |||
Table 1: The composition of the solutions.
