In situ Ca2+ imaging of the PKJ can reveal important cellular activity of RP pacemaker cells. By using mice that express genetically encoded Ca2+ indicators (such as GCaMP), driven by cell-specific promotors, information about RP pacemaking can be obtained with accuracy and detail that is not possible from Ca2+ imaging experiments from flat-sheet RP preparations. The beginning of the PKJ is distinguished by the sudden appearance of semi-circles of muscle suspended between kidney parenchymal tissue (Figure 2C; proximal PKJ enclosed in dashed box). During subsequent rounds of sectioning, the inner medulla becomes distinguishable from the surrounding cortical tissue. Under a light microscope, the inner medulla appears striated in regions, lighter in color compared to cortical tissue and discontinuous on its long axis with the rest of the kidney (Figure 2B,D). At this point, more PKJ regions will start to appear. Examples of this are shown in Figure 2D (dashed rectangles, H and G) where 3 semi-circles of muscle are suspended by parenchymal tissue. These muscle bands will be closely apposed to the inner papilla and will typically neighbor a renal arteriole (Figure 2D, dashed rectangles; Figure 2F–H, black arrowheads). As more distal sections are derived, these bands of muscle will integrate to form a more complete, unified structure, indicating the end of the PKJ region (Figure 2E).
Figure 3A,B shows a PKJ section at low power (4-10x) from a mouse expressing GCaMP in PDGFRα+ cells (GCaMP6f expressed by inducible Cre-recombinase driven by Pdgfra). Using landmarks such as the renal arteriole (Figure 3A; asterisk), experimenters should be able to readily distinguish the thin PKJ wall suspended between parenchymal tissue (Figure 3B; asterisks). The expression of GCaMP6f in this specific transgenic tissue is spread across the entire width of the PKJ, across both the muscle and adventitial layers (Figure 3C).
In PDGFRα+ GCaMP6f+ kidney slices, a network of cells that typically extends over the width of the PKJ wall will be fluorescent (Figure 3C) and display oscillating Ca2+ transients of various durations and frequencies. PDGFRα+ cells in the PKJ wall display two different types of Ca2+ transient durations. In the adventitial layer (orientated closer to cortex), PDGFRα+ cells present as a network of cells and their processes are defined. Adventitial PDGFRα+ cells exhibit low-frequency (4 ± 2.7 Hz) and long-duration (1 ± 0.67 s) Ca2+ transients. The second layer of PDGFRα+ cells, present in the muscle layer (orientated closer to the medulla), exhibit similar Ca2+ transient frequencies and durations as SMC GCaMP3+ cells (described below) as they are the same cell type.
In SMC GCaMP3+ kidney slices, a layer of GCaMP3+ cells is present in the muscle layer (Figure 3D). There will be no fluorescent signal in the adventitial layer (Figure 3D; asterisk). GCaMP3+ SMCs in the muscular layer typically exhibit high-frequency (10 ± 4 Hz) and short-duration (632 ± 74 s) Ca2+ transients. PDGFRα+ cells located in the PKJ adventitia elicit long-duration, low-frequency Ca2+ transients (Figure 3E, Video 1). However, Ca2+ imaging experiments from tissue expressing GCaMP3 driven by the Myh11 promotor is restricted to the muscular aspect of the PKJ (Figure 3D). Compared to PDGFRα+ cells in the adventitia, SMCs fired shorter duration Ca2+ transients more frequently (Figure 3F, Video 2).
In addition to understanding signaling properties in PKJ PDGFRα+ ICs, the application of this technique to study other cell types in the vibratome-sectioned kidney has been demonstrated in this paper. Upon close examination of the renal medulla (in mice expressing GCaMP6f in PDGFRα+ cells), an array of fluorescent Ca2+ signals within and surrounding collecting ducts (Video 3) was observed. Medullary PDGFRα+ cells fired spontaneous Ca2+ transients of variable frequency and duration. These Ca2+ imaging studies of kidney vibratome sections could also be expanded to studying renal arterioles (~50-80 mm diameter) that often neighbor PKJ muscle segments (Figure 2F,G; white arrows). Ca2+ imaging of renal arterioles (from tissue expressing GCaMP in smooth muscle cells) demonstrates oscillating Ca2+ transients in SMCs (Video 4).
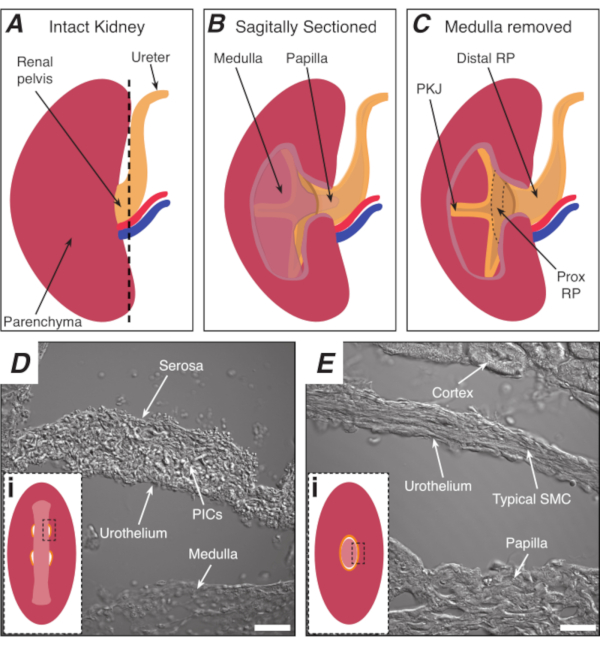
Figure 1: Basic kidney anatomy and location of PKJ pacemaker region. (A) Diagram of the intact kidney showing the orientation of the RP and ureter. The renal artery and renal vein are displayed in red and blue, respectively. (B) The intact kidney can be cut along a sagittal plane to expose the inner aspect of the kidney, including the medulla, papilla (distal medulla where collecting ducts converge), and proximal and distal RP. (C) The medulla and papilla can be excised to completely expose the PKJ and prox RP. (D and E) represent transmitted light images from the PKJ pacemaker region and distal RP, respectively. Sequential sectioning towards the distal end of the pelvis results in the semicircles of muscle in the PKJ region (Di) combining into one, thicker muscular ring (Ei) that encapsulates the entire papilla. Black, dashed rectangles in Di and Ei show approximate areas in coronal kidney sections where transmitted light images were acquired. Orientation of images D and E are 90° anti-clockwise to respective insets (Di and Ei). Scale bars in D and E = 20 µm. Abbreviations: RP = renal pelvis; prox RP = proximal renal pelvis; PKJ = pelvic-kidney junction; PICs = platelet-derived growth factor receptor-alpha-positive interstitial cells; SMC = smooth muscle cell. Please click here to view a larger version of this figure.
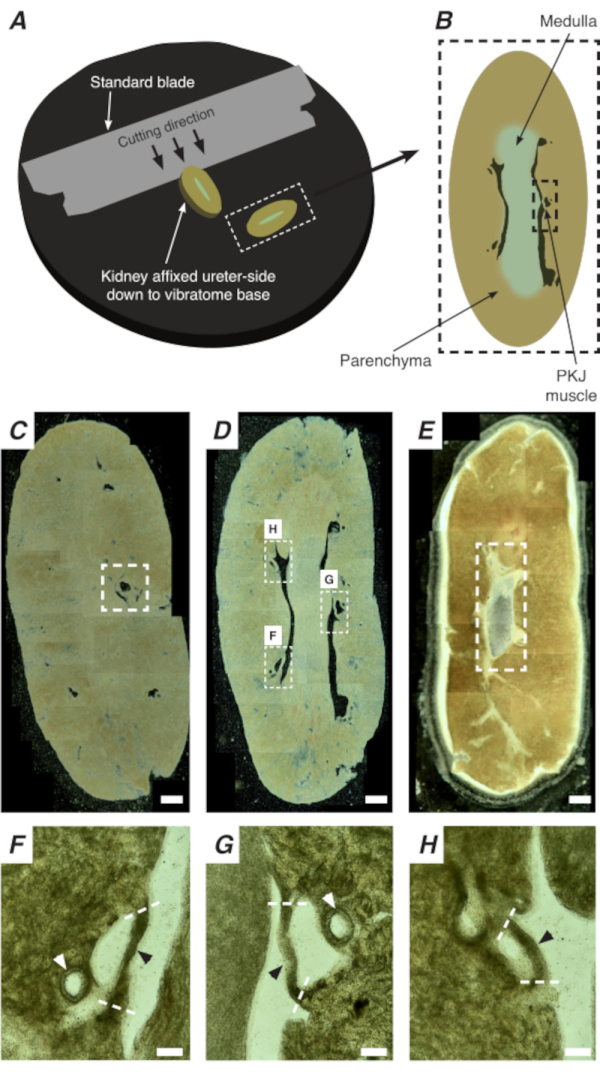
Figure 2: Vibratome sectioning of whole kidneys to generate thin sections. (A) Kidneys are mounted ureter side down to the base of the vibratome instrument, and a standard blade (attached to the vibratome head) is used to cut sequential sections from the proximal to distal end of the kidney. (B) Diagrammatic representation of a thin section cut from the whole kidney with annotated landmarks. PKJ muscle segments (black dashed rectangle) are often found suspended between parenchymal tissue. (C) Light microscopic image of a proximal kidney section. The appearance of muscle bands suspended between parenchymal tissue indicates the beginning of the proximal PKJ projections (indicated inside white dashed rectangle). (D) Light microscopic image representing the optimal region where multiple (2-3) PKJ segments can be found (areas within white dashed rectangles). Thin PKJ muscle strips are suspended between the kidney parenchyma and align closely with renal arterioles and medulla. (E) Light microscopic image of a distal kidney section. Individual muscle segments have merged to form a single, continuous muscle band (white dashed rectangle) that surrounds the inner papilla (not present in this image). Scale bars C–E = 500 µm. F–H Zoomed (20x) regions from panel D indicate the location of the PKJ (black arrowheads), renal arterioles (white arrowheads), and cut sites for isolating the PKJ (dashed white lines). Scale bars F–H = 100 µm. Abbreviation: PKJ = pelvic-kidney junction. Please click here to view a larger version of this figure.
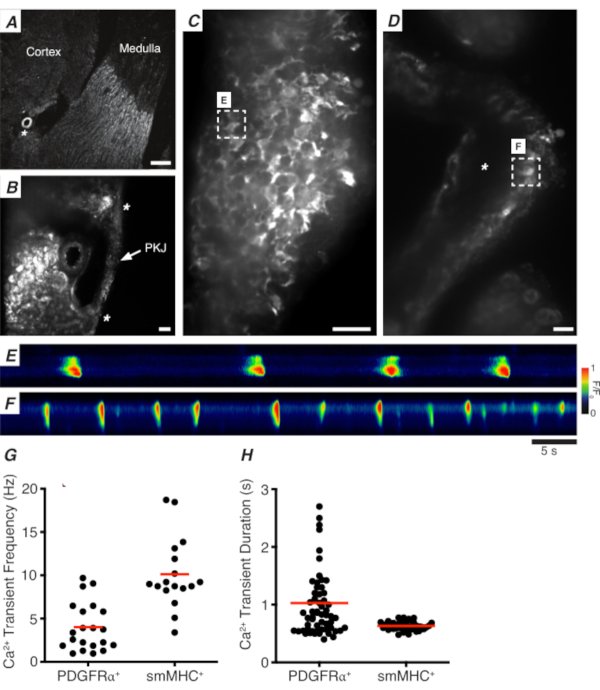
Figure 3: Ca2+ imaging of vibratome sections. (A) Representative low-power image (4x) of a vibratome section denoting location of the renal arteriole (asterisk). Scale bar = 200 µm. (B) Zoomed (20x) representative image of the PKJ (labeled) suspended between kidney parenchymal tissue denoting locations of the PKJ muscle (white arrowhead), renal arteriole (asterisk). Scale bar = 50 µm. (C) High-power (40x) image of the PKJ expressing GCaMP in PDGFRα+ cells. Scale bar = 20 µm. (D) High-power (20x) image of the PKJ expressing GCaMP in smooth muscle cells. Scale bar = 20 µm. (E) Spatiotemporal map of Ca2+ transients sampled from a GCaMP+ PDGFRα+ cell indicated in panel C. Look up table coded for F/F0. (F) Spatiotemporal map of Ca2+ transients sampled from a GCaMP+ PDGFRα+ cell indicated in panel D. Look up table coded for F/F0. (G) Representative data for Ca2+ transient frequency (Hz) for GCaMP+ PDGFRα+ cells and GCaMP+ smMHC cells. (H) Representative data for Ca2+ transient duration (s) for GCaMP+ PDGFRα+ cells and GCaMP+ smMHC cells. Abbreviations: PKJ = pelvic-kidney junction; PDGFRα+ = platelet-derived growth factor receptor-alpha-positive; smMHC = smooth muscle myosin heavy chain. Please click here to view a larger version of this figure.
Video 1: Spontaneous Ca2+ transients in GCaMP+ PDGFRα+ cells in PKJ vibratome sections. Abbreviations: PKJ = pelvic-kidney junction; PDGFRα+ = platelet-derived growth factor receptor-alpha-positive. Please click here to view this video. (Right-click to download.)
Video 2: Spontaneous Ca2+ transients in GCaMP+ smooth muscle cells in pelvic-kidney junction vibratome sections. Please click here to view this video. (Right-click to download.)
Video 3: Spontaneous Ca2+ transients in GCaMP+ PDGFRα+ cells in renal medullary vibratome sections. Abbreviation: PDGFRα+ = platelet-derived growth factor receptor-alpha-positive. Please click here to view this video. (Right-click to download.)
Video 4: Low-amplitude Ca2+ transient activity in the vibratome sections of the renal arteriole. Please click here to view this video. (Right-click to download.)
