Ubiquitous in vivo RNA Targeting Using a Two-Component CasRx System
The F1 transheterozygous flies expressing both the Ubiq-CasRx and the gRNA (targeting both endogenous and exogenous genes) constructs showed marked phenotypes compared to the control flies expressing the Ubiq-dCasRx and gRNA constructs (Figure 2 and Figure 4). Specifically, the transheterozygous CasRx flies have significantly lower levels of survival rate compared to the transheterozgyous dCasRx flies, indicating toxicity of the Ubiq-CasRx system (Figure 2A and Figure 4A). It is worth noting that both transheterozygous CasRx and dCasRx flies have less than 50% inheritance rate, which is the expected ratio based on Mendelian genetics. Of the three target genes, the Ubiq-CasRx/+; U6-gRNAN/+ flies and Ubiq-CasRx/+; U6-gRNAy/+ flies are non-viable (0% inheritance) and did not grow beyond the second instar larvae stage (Figure 2A-2B). The surviving Ubiq-CasRx/+; U6-gRNAw/+ flies, the inheritance of which was 12.9%, showed a distinct fully-penetrant white-eyed phenotype (Figure 2B). In addition to observable traits associated with CasRx, we were able to confirm significant reduction of target gene transcripts for 3 target genes: Notch, yellow, and GFP (Figure 2E-2G). Reduction of white gene transcripts was observed in Ubiq-CasRx/+, U6-gRNAw/+ flies, compared to the control Ubiq-dCasRx/+, U6-gRNAw/+ flies, though the reduction was not statistically significant (Figure 2E – 2F). Evidence of off-target activity induced by CasRx was found when comparing the differentially expressed transcripts between samples from CasRx-expressing flies and samples from dCasRx-expressing flies (Figure 2E, 2G). The number of non-target transcripts significantly differentially expressed are as follows: white, 253 (1.4% of total transcripts); Notch, 300 (1.7%); yellow, 41 (0.23%); GFP, 5,880 (33%) (Figure2G). Out of the total 17,779 different transcripts, 6 non-target transcripts were significantly differentially expressed in all 4 groups of samples. One of the 6 transcripts identified was Gadd45, a gene involved in apoptosis and cellular arrest in flies, raising the possibility that the enzymatic action of CasRx may either directly trigger cellular apoptosis or indirectly trigger misexpression of other genes, which in turn leads to apoptosis. Finally, it is worth noting that the Ubiq-CasRx and Ubiq-dCasRx flies were not established as homozygous stocks, presumably due to toxicity conferred by high ubiquitous expression. As a result, heterozygous Ubiq-CasRx/CyO and Ubiq-dCasRx/CyO flies were used for crossing with homozygous gRNA fly lines. In sum, the two-component Ubiq-CasRx system is able to achieve ubiquitous RNA targeting for both endogenous and exogenous targets resulting in observable phenotypes and transcript reduction. These results also showed that CasRx-mediated RNA targeting may introduce toxicity in vivo.
Ubiquitous in vivo exogenous RNA Targeting Using a Three-Component CasRx System
The results from the two-step cross showed that despite the exogenous nature of the target gene (i.e., Fluc), expressing all three transgenes in F2 triple transheterozygotes (Ubiq-CasRx/+; gRNAFluc/Ubiq-Fluc-Ubiq-Rluc) resulted in 100% lethality compared to control crosses involving Ubiq-dCasRx, where no lethality was observed in the F2 triple transheterozygotes (Ubiq-dCasRx/+; gRNAFluc/Ubiq-Fluc-Ubiq-Rluc) (Figure 3B-C). More specifically, only the combination of all three transgenes (Ubiq-CasRx/+; gRNAFluc/Ubiq-Fluc-Ubiq-Rluc) resulted in 100% lethality (Figure 3B and D), while (Ubiq-CasRx/+; gRNAFluc/TM6) and (Ubiq-CasRx/+; Ubiq-Fluc-Ubiq-Rluc/TM6) genotypes were viable and lacked phenotypes with their inheritance rates matching the expected Mendelian transmission rates, suggesting that the availability of the target sequence (i.e., firefly luciferase) in combination with Ubiq-CasRx/+ and the gRNAFluc is what resulted in the observed lethality phenotypes, presumably stemming from the collateral activity of Cas13 enzymes2,8. In addition, no distinguishable phenotypes or dramatic influence on inheritance in F1 transheterozygotes (Ubiq-CasRx/+; gRNAFluc/+ or Ubiq-CasRx/+; Ubiq-Fluc-Ubiq-Rluc/+) were observed compared to Ubiq-dCasRx controls (Ubiq-dCasRx/+; gRNAFluc/+ or Ubiq-dCasRx/+; Ubiq-Fluc-Ubiq-Rluc/+) (Figure 3B), indicating that a catalytically active enzyme is essential to obtain the lethality phenotypes observed. Furthermore, Fluc and Rluc expression levels in flies of all viable genotypes did not show significant reduction in Fluc expression in the Ubiq-dCasRx triple transheterozygotes (Ubiq-dCasRx/+; gRNAFluc/Ubiq-Fluc-Ubiq-Rluc) compared to dual luciferase reporter controls. This suggests that Fluc protein expression levels were not reduced by dCasRx targeting (Figure 3D). Taken together, the common lethality phenotype in the two different CasRx-mediated ubiquitous RNA targeting experiments indicate that when used on tissues ubiquitously, CasRx-mediated RNA targeting can be toxic to the organism.
Tissue-specific in vivo RNA Targeting Using a Three-Component CasRx System
The high level of toxicity observed in ubiquitous RNA targeting experiments prompted us to explore the tissue-specific RNA targeting using a three-component CasRx system design detailed in the methods section. Indeed, the level of toxicity observed was reduced when the overall CasRx expression was lowered using the UASt promoter compared to that of the Ubiq promoter, this is exemplified in three aspects: 1) the UASt-CasRx and UASt-dCasRx lines were maintained as homozygous lines, though based on the two-step cross scheme double balanced UASt-CasRx and UASt-dCasRx lines were used to perform the crosses, 2) all F2 generation dCasRx triple transheterozygous inheritance rates matched the expected 25% Mendelian inheritance rate, and 3) the F2 generation CasRx triple transheterozygous lethality phenotype was moderately reduced. In the white targeting experiment, of the 25% Mendelian inheritance rates expected in the F2 triple transheterozygotes, only 0.57% viable adult flies (UASt-CasRx/+; gRNAw/GMR-Gal4) were observed, all of which displayed severe eye specific pigmentation and morphology phenotypes (Figure 4A and 4B). For the white-targeting cross, the CasRx-expressing triple transheterozygous F2 inheritance rate was significantly lower than that of the dCasRx-expressing triple transheterozygous control group (27.6%) (Figure 4A). In the Notch targeting experiment, CasRx-expressing triple tranheterozygous carrying all three transgenes were 100% lethal, while the dCasRx control inheritance rate was 29.3% (Figure 4A). In the yellow targeting experiment, F2 triple transheterozygous CasRx-expressing, gRNAy, and y-GAL4 showed marginal chitin pigment reduction as small patches of yellow cuticle on the thorax and abdomen with an inheritance rate of 2.67%, much lower than that of the dCasRx control group (25.2%) (Figure4A). All dCasRx control triple transheterozygous flies did not present obvious phenotypes as the CasRx-expressing flies, indicating that catalytic activity of CasRx contributed to the phenotypes observed. The low inheritance rate in the CasRx triple transheterozygous group suggested that two sources of toxicity exist in CasRx RNA targeting: one is associated with high expression of CasRx, the toxicity of which was reduced by restrictive CasRx expression, the other is associated with the collateral activity. Taken together, these results showed that the CasRx system can achieve tissue-specific in vivo RNA targeting by leveraging the classical Gal4/UASt system and in the meantime reduce the toxicity. However, toxicity and occasional lethality phenotypes were still observed at a lower level of severity compared to that of the ubiquitous approaches, indicating that collateral cleavage activity is associated with toxicity.
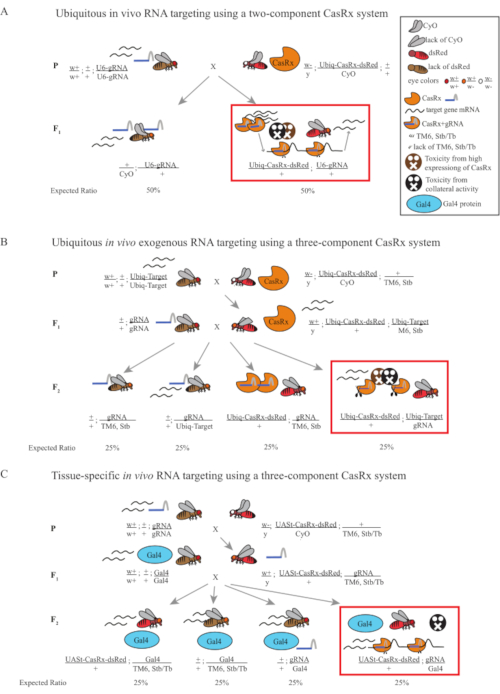
Figure 1: General overview of RNA targeting using a Cas13D system. (A) Schematics of the one-step genetic cross in the ubiquitous in vivo RNA targeting using a two-component CasRx system. (B) Schematics of a two-step genetic cross in the ubiquitous in vivo exogenous RNA targeting using the three-component CasRx system. (C) Schematics of a two-step genetic cross in the tissue-specific in vivo RNA targeting using a three-component CasRx system. Please click here to view a larger version of this figure.
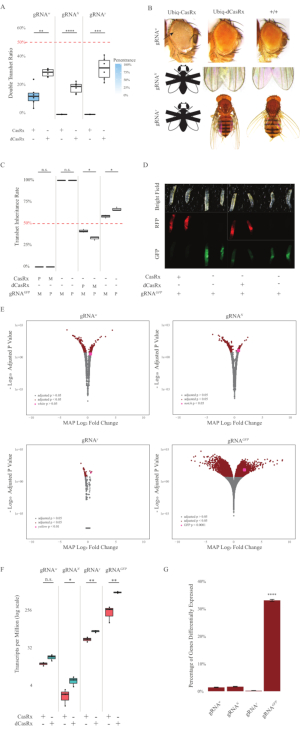
Figure 2: Ubiquitous in vivo RNA targeting using a two-component CasRx system (reprinted5). (A) Total inheritance percentages of transheterozygous flies inheriting Ubiq-CasRx (or Ubiq-dCasRx) and gRNAs. Blue shading in the box plot indicates phenotype penetrance. (B) Phenotypes of transheterozygous flies. Arrows indicate tissue necrosis in the eye. Black and white fly marked with ''X'' represents lethality. (C) Total inheritance percentages of transheterozygous flies of bidirectional crosses between Ubiq-CasRx (or Ubiq-dCasRx) and gRNAGFP-OpIE2-GFP flies. M, maternal inheritance of CasRx; P, paternal inheritance of CasRx. (D) F1 larvae progenies in the paternal cross. (E) Transcripts' maximum a posteriori estimates for the logarithmic fold change. DESeq2 pipeline was used. (F) Transcripts per million (TPM) targeted with CasRx or dCasRx. (G) CasRx-depentent differentially expressed transcript percentage of transcripts. Please click here to view a larger version of this figure.
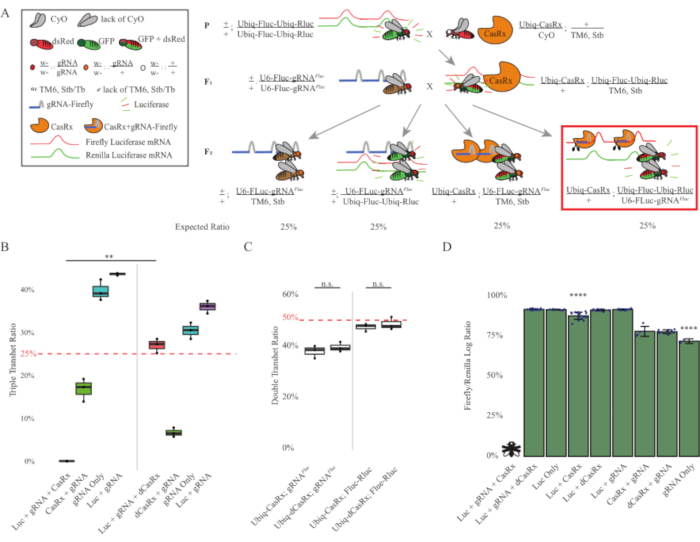
Figure 3: Ubiquitous in vivo exogenous RNA targeting using a three-component CasRx system. (A) Schematics of the two-step genetic cross. (B) Total inheritance percentages for all genotypes emerging in the F2 generation. Inheriting all three transgenes (Ubiq-CasRx, Ubiq-Fluc-Ubiq-Rluc, and gRNAFLuc) in F2 progeny resulted in 100% lethality and was significantly lower compared to the Ubiq-dCasRx triple transheterozygotes control group (p = 0.001, t-test). (C) Carrying Ubiq-CasRx/gRNAFluc alone or Ubiq-CasRx and Ubiq-Fluc-Ubiq-Rluc alone did not lead to severe lethality, and inheritance ratios between Ubiq-CasRx and Ubiq-dCasRx transheterozygotes were not significantly different (p = 0.41 and p = 0.51, respectively, t-test). (D) Luciferase ratios normalizing Fluc readings to Rluc readings. Triple transheterozygous flies expressing Ubiq-CasRx, Ubiq-Fluc-Ubiq-Rluc, gRNAFLuc were embryonic lethal, which was represented by a fly with an "X", and as a result luciferase expression was not measured. Fluc/Rluc ratio of Ubiq-CasRx/+, Ubiq-Fluc-Ubiq-Rluc/TM6, Stb transheterozygotes was significantly lower than that of the other Ubiq-Fluc-Ubiq-Rluc-expressing groups (p = 1.2e-06 or lower, t-test). The results from the gRNAFLuc-only group were significantly lower than that of all other groups (p = 1.2e-06 or lower, t-test). Please click here to view a larger version of this figure.
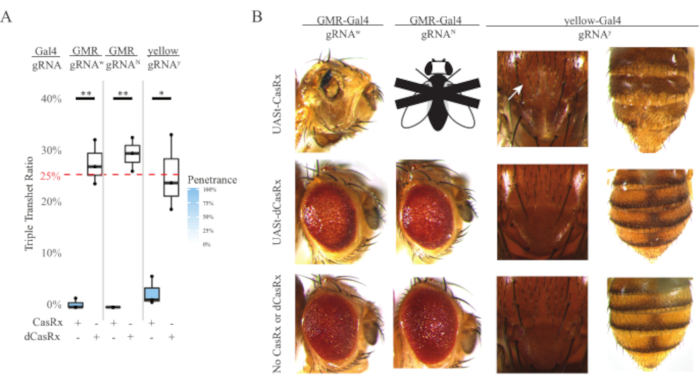
Figure 4: Tissue-specific in vivo RNA targeting using a three-component CasRx system (reprinted5). (A) Total inheritance percentage of triple transheterozygous flies carrying three transgenes (UASt-CasRx or UASt-dCasRx, gRNAs, and Gal4-driver. (B) Phenotypes of the triple transheterozygous flies. The white arrow indicates chitin pigment reduction in the thorax. Black and white fly marked with ''X'' represents lethality. Please click here to view a larger version of this figure.
| Construct | Description | Primer | Primer Sequence (5’ to 3’) | PCR Template |
| OA-1050E | CasRx | 1050E.C3 | TACTAATTTTCCAC ATCTCTATTTTGAC CCGCAGATTAATTA ATGAGCCCCAAGA AGAA |
pNLS-RfxCas13d-NLS-HA (pCasRx) |
| 1050E.C4 | CAATTGATTTGTTA TTTTAAAAACGATT CATTCTAGCTAGCT TAAGCGTAATCTGG AACA |
|||
| OA-1050R | dCasRx | 1050E.C3 | TACTAATTTTCCAC ATCTCTATTTTGAC CCGCAGATTAATTA ATGAGCCCCAAGA AGAA |
pNLS-dRfxCas13d-NLS-HA (pdCasRx) |
| 1050E.C4 | CAATTGATTTGTTA TTTTAAAAACGATT CATTCTAGCTAGCT TAAGCGTAATCTG GAACA |
|||
| OA-1050L | UASt promoter | 1041.C9 | GCGGGTTCTCGA CGGTCACGGCGG GCATGTCGACGC GGCCGCAACCAA CAACACTAGTAG |
pJFRC81 |
| 1041.C11 | CTGGCCTCCACC TTTCTCTTCTTCT TGGGGCTCATGT TTAAACCCAATT CCCTATTCAGA |
|||
| CasRx | 1050L.C1 | AATACAAGAAGA GAACTCTGAATA GGGAATTGGGT TTAAACATGAGC CCCAAGAAGAA |
pCasRx | |
| 1050E.C4 | CAATTGATTTGT TATTTTAAAAAC GATTCATTCTA GCTAGCTTAAG CGTAATCTGGA ACA |
|||
| OA-1050S | UASt promoter | 1041.C9 | GCGGGTTCTC GACGGTCACG GCGGGCATGT CGACGCGGCC GCAACCAACAA CACTAGTAG |
pJFRC81 |
| 1041.C11 | CTGGCCTCCA CCTTTCTCTTC TTCTTGGGGCT CATGTTTAAAC CCAATTCCCTA TTCAGA |
|||
| dCasRx | 1050L.C1 | AATACAAGAAG AGAACTCTGAAT AGGGAATTGGG TTTAAACATGAG CCCCAAGAAGAA |
pdCasRx | |
| 1050E.C4 | CAATTGATTTGT TATTTTAAAAAC GATTCATTCTAG CTAGCTTAAGCG TAATCTGGAACA |
|||
| OA-1043 | U6:3 promoter | 1043.C1 | GGGAATTGGGA ATTGGGCAATAT TTAAATGGCGGC GCGCCGAATTCT TTTTTGCTCACCT |
Addgene plasmid #164586 |
| 1043.C23 | ACACTAGTGGAT CTCTAGAGGTAC CGTTGCGGCCG CAAAAAAGTTGT AATAGCCCCTCA AAACTGGACCTT CCACAACTGCAG CCGACGTTAAAT TGAAA |
|||
| OA-1052B | Ubiq promoter | 1052B.C1 | GGGAATTGGGCA ATATTTAAATGGC GGCTGCAGCGC GCAGATCGCCGAT |
Addgene plasmid #112686 |
| 1052B.C2 | TTTCTTTATGTTT TTGGCGTCTTCC ATCCTAGGTCTG CGGGTCAAAATA GAGATG |
|||
| T2A-eGFP | 908A1 | ATAAAGGCCAAG AAGGGCGGAAA GATCGCCGTGG AGGGCAGAGGA AGTCTTCTAACAT GC |
Addgene plasmid #112686 | |
| 908A2 | TTGTTATTTTAAAA ACGATTCATTCTA GGCGATCGCTTA CTTGTACAGCTC GTCCATGCC |
|||
| Reversed Ubiq promoter | 908A3 | ACCGTGACCTAC ATCGTCGACACTA GTGGATCTCTAGA CGCGCAGATCGC CGATG |
Addgene plasmid #112686 | |
| 908A4 | GGATCATAAACTT TCGAAGTCATGC GGCCGCTCTGCG GGTCAAAATAGAG ATGT |
Table 1: List of molecular construct and primers used in this study. This list includes all constructs (both the ID and description) and each construct's associated primers (both the ID and sequences (5' to 3')) and templates used.
