This technique allows the investigator to non-invasively transplant iPSMG into the hippocampus and cerebellum but not into the cortex of the mouse brain. After completing the study, anesthetized mice were subjected to transcardial perfusion of ice-cold PBS(−), followed by ice-cold 4% (w/v) paraformaldehyde in PBS. The brains were isolated, postfixed overnight in 4% (w/v) paraformaldehyde, and cryoprotected in a PBS containing 30% (w/v) sucrose. Further, the brains were frozen in an embedding compound and sectioned (20 µm thick coronal sections) on a cryostat. The sections were washed thrice in PBS(−) (10 min each) and permeabilized and blocked with 0.5% (v/v) Triton X-100 in 10% normal goat serum for 1 h. The sections were then incubated with anti-human-specific cytoplasm marker, STEM121 (1:100), and anti Iba1 (1:1000) for 5 days. Next, the sections were washed three times with PBS(−) for 10 min each and were incubated with secondary antibodies: Alexa Fluor 488-, or 546-conjugated mouse, or rabbit IgGs (1:1000) for 2 h at room temperature. After washing with PBS(-) three times, sections were mounted on slides using an antifade mounting medium. A confocal microscope equipped with a 40x objective lens was used for acquiring fluorescence images. The viability of iPSMG at 2 months post-transplantation in the hippocampus and the cortex are shown in Figure 1. The number of transplanted cells can be determined by counting cells that are positive for both human-specific antibodies and pan-microglial/monocyte markers, whereas endogenous mouse microglia are positive for pan-microglial/monocyte marker only as previously described18. The transplanted iPSMGs replace mouse microglia, show ramified morphologies in the hippocampus, and are not detected in the cortex18.
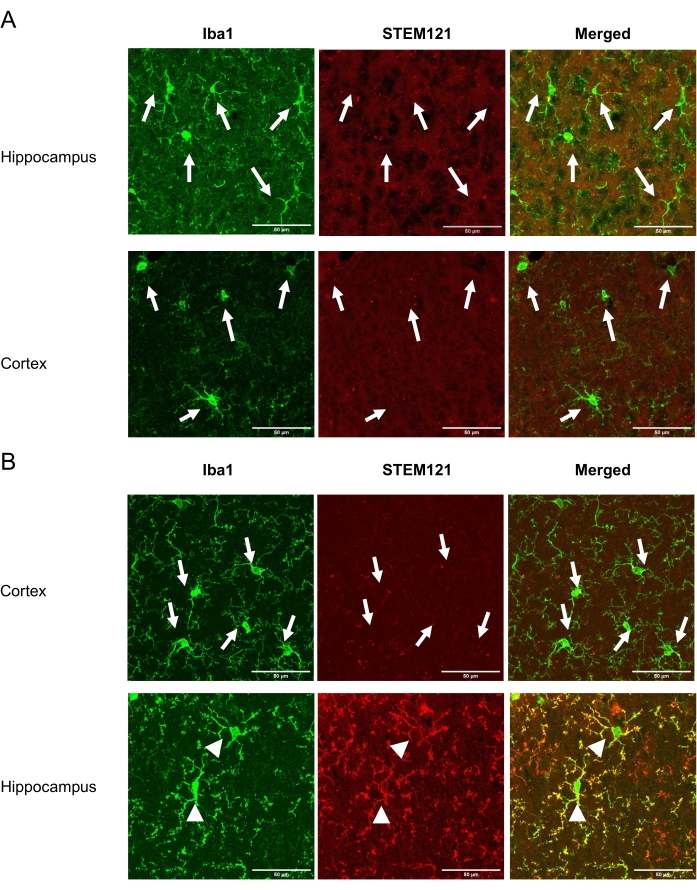
Figure 1: Viability of iPSMG in the cortex and the hippocampus at 2 months after Tsn. The left panel shows immunostaining with a pan-microglial/monocyte marker, Iba1 (green). The middle panel shows immunostaining with human-specific cytoplasm marker STEM121 (red). The right panel shows a merged image of Iba1 and STEM121 immunostaining. (A) In control mice, only mouse microglia (Iba1+/STEM121-) were detected in both cortex and hippocampus. (B) In iPSMG transplanted mice, in the cortex, only mouse microglia (Iba1+/STEM121–) were detected, whereas in the hippocampus iPSMG (Iba1+/STEM121+) were detected. The arrows in the image show mouse microglia, whereas arrowheads show iPSMG. A confocal microscope equipped with a 40x objective lens was used for fluorescence image acquisition. Maximum image size: 1024 x 1024 pixels. Zoom factor: 2. Scale bars = 50 µm Please click here to view a larger version of this figure.
