Following the detailed protocol herein for the preparation of supported lipid bilayers (SLBs) in flow chambers (Figure 1), one can clearly recognize a speckle-like pattern in the native view of all displayed conditions (Figure 2). This effect is caused by the surface roughness of the glass, which generally dominates the scattering signal and leads to visually indistinguishable conditions (glass, glass with SLB, or glass with SLB and attached proteins). The presence of vesicles, however, is clearly distinct due to the vesicles' large scattering cross-section and enables the observation of vesicle rupture and fusion into homogeneous membranes (Figure 2B and Supplementary Movie 1). When removing the static scattering signal of the glass surface with a ratiometric approach that emphasizes the dynamic elements within the field of view24,25, one can uncover unlabeled proteins diffusing on the membrane (Figure 2D) while an empty SLB (Figure 2C) or glass itself (Figure 2A) appears as a noisy image.
The inherent background of MSPT measurements can be locally estimated by dividing each pixel value through the median of n preceding and succeeding pixels of the movie at the same image position (Figure 3). As a result, macromolecules appear as isotropic point-spread functions (PSFs) whose motion on the membrane can be observed, tracked, and quantified. In fact, the availability of both contrast and dynamic behavior enables the direct relation of a particle's molecular size to its respective diffusive behavior, all without the need for labeling the particle. Nevertheless, to interpret the iSCAT contrast determined during MSPT experiments, it is essential to perform a calibration that translates the signal amplitude into molecular mass. This can be achieved by attaching biomolecules of known mass to an SLB via a biotin-streptavidin-biotin complex (Figure 4A). As an exemplary strategy, one can use biotinylated variants of bovine serum albumin (BSA), protein A (prA), alkaline phosphatase (AP), and fibronectin (FN), which bind to streptavidin (STP) that itself is bound to biotin-containing lipids (Biotinyl Cap PE) in the membrane. As displayed in Figure 4A, the increasingly pronounced contrast of these exemplary macromolecules reflects the increasing molecular weight of the respective biotinylated standards. By assigning each peak of the contrast histograms (Figure 4B) to the corresponding mass of the standard protein's oligomer state, a linear relationship between contrast and mass is revealed21,22 and can subsequently be used for the analysis of unknown macromolecule systems (Figure 4C).
A good example demonstrating the applicability and capabilities of MSPT to analyze molecular weights and hence study oligomer states and oligomerization events is the consideration of biotinylated aldolase and biotinylated IgG (Figure 5). Aldolase is commonly reported to be a homotetramer32. However, the mass distribution resolved by MSPT features four distinct peaks, which highlights the presence of multiple populations (Figure 5A). While the first minor peak corresponds to unoccupied streptavidin and can be expected due to the configuration in this kind of experiment, aldolase complexes with only two subunits (2SU) or six subunits (6SU) can likewise be detected (Figure 5B). Interestingly, tetra- and hexameric aldolase-streptavidin complexes display a reduced diffusion coefficient when compared to dimeric aldolase and streptavidin alone, indicating an increased viscous drag, e.g., via the attachment of a second biotinylated lipid to the streptavidin. Similarly, biotinylated IgG exhibits three peaks in the mass distribution, with the first peak again matching the mass of a single streptavidin. The mass of the most abundant peak corresponds to the mass of one light and one heavy chain (1SU), i.e., one-half of an IgG antibody. The full antibody with two identical halves (2SU) is detected in about 11% of the cases. The decrease of the diffusion coefficient with increasing complex sizes indicates interactions of the streptavidin with more than one biotinylated lipid or additional drag caused by the attached IgG, or both.
Besides the sole analysis of membrane-dependent oligomer states, MSPT also confers the particular advantage of correlating the diffusive behavior of a macromolecule of interest with its oligomer state. Representative results for this type of analysis are shown for annexin V (AnV) and cholera toxin subunit B (CTxB), which bind to dioleoylphosphatidylserine (DOPS) or glycosphingolipids (GM1), respectively, incorporated in the membrane (Figure 6A). Both kernel density estimations (KDEs) feature unimodal distributions of mass and diffusion, indicating a single abundant species with similar diffusive behavior. The peak position of molecular mass and diffusion coefficient was found to be 49.8 ± 2.2 kDa and 1.4 ± 0.1 µm2/s, respectively, for AnV as well as 62.7 ± 3.1 kDa and 0.4 ± 0.1 µm2/s, respectively, for CTxB. The measured diffusion coefficients are comparable to previously reported values obtained from high-speed AFM and FRAP33,34. The slightly reduced mass as compared to the mass of the expected macromolecule (52 kDa for an AnV trimer, 65 kDa for a CTxB pentamer) may indicate the presence of smaller complexes with fewer subunits in the ensemble. While the mass difference between the proteins is small and close to the specified detection limit of the microscope (≈50 kDa), their diffusion coefficients differ considerably. In an equimolar mixture, for instance, by comparing the diffusion of the mixture to the distribution of AnV and CTxB alone, one can conclude that AnV is more abundant on the membrane than CTxB (Figure 6B). However, if the concentration of CTxB is doubled as compared to the concentration of AnV, the equilibrium is shifted toward CTxB as the predominant protein on the membrane. As illustrated for mixtures of AnV and CTxB, MSPT not only allows to discriminate membrane-associated macromolecules according to their molecular weight but also enables the discrimination of different macromolecule populations according to their diffusive behavior.
As with all microscopy techniques, some experimental requirements are crucial in order to achieve the desired quality of data. An important example in this context is thoroughly cleaned coverslips. In general, this is considered a prerequisite for microscopy-related single-molecule experiments, but MSPT is particularly sensitive to sample impurities. The increased scattering originating from the glass surface of uncleaned coverslips prevents any quantitative iSCAT measurement. Notably, even residual dirt or dust particles on insufficiently cleaned glass can cause notable image distortions, recognizable as bright spots in the native imaging mode (Figure 7A). Although these defects are removed by the background estimation due to their static nature, accurate determination of a particle's contrast might be impaired and thus negatively influences its quantitative analysis. Another common problem encountered in MSPT experiments are the remaining vesicles that either float (encircled in orange) through the field of view or unfused vesicles that are stuck (encircled in blue) at a specific position on the membrane and appear as large pulsating scatterers (Figure 7B). To minimize their occurrence and interference with movie acquisition, it is recommended to thoroughly wash the SLB before adding the protein and to use freshly prepared mixtures of small unilamellar vesicles (SUVs) and divalent cations.
One factor that must be taken into account for the design of mass-sensitive particle tracking experiments is the density of macromolecules associated with the membrane interface. High particle densities on the membrane can in fact cause problems for two reasons: i) The linking of particle detections from consecutive frames into trajectories becomes ambiguous and hence increases the likelihood of errors and misjudged diffusion coefficients. ii) The mass of particles, which is extracted from the amplitude of their corresponding PSF fit, becomes systematically underestimated and mass peaks broaden because the separation of the static background signal from the dynamic particle signal is increasingly difficult (Figure 7C). Currently, visual assessment of data quality in the process of acquiring MSPT videos is difficult on the available commercial microscopes because the implemented ratiometric view in the acquisition software is using the background removal established for mass photometry21 instead of the median-based algorithm described here and in references24,25 (Figure 7D). The mean-based continuous background removal used to visualize landing molecules in mass photometry causes diffusing particles to appear as dark fronts with bright tails, which makes the spots appear highly anisotropic and interferes with PSF fitting during the detection procedure. Thus, the use of the implemented mean-based image processing in the acquisition software is unsuitable for the analysis of diffusing biomolecules on membranes.
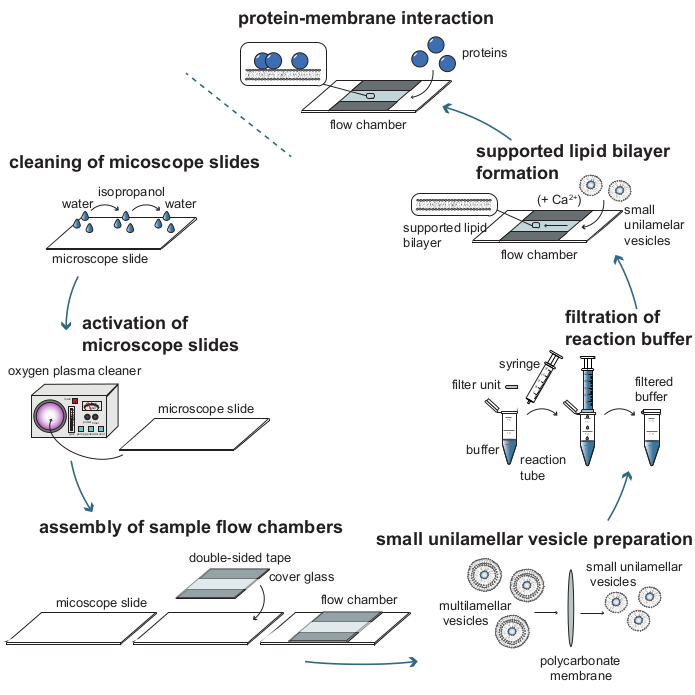
Figure 1: Process flow diagram of the individual steps required to analyze protein-membrane interactions with mass-sensitive particle tracking (MSPT). To prepare samples for MSPT measurements, glass cover slides must be thoroughly cleaned and activated with an oxygen plasma. After their assembly into sample flow chambers, small unilamellar vesicles (SUVs) are prepared for supported lipid bilayer (SLB) formation and all reaction buffers are filtered to reduce background scattering. SUVs are added to form lipid bilayers in the flow chambers. Optionally, divalent cations such as Ca2+ ions may be added to the SUVs to promote vesicle rupture. At last, low concentrations of the protein of interest are flushed into the reaction chamber. Please click here to view a larger version of this figure.
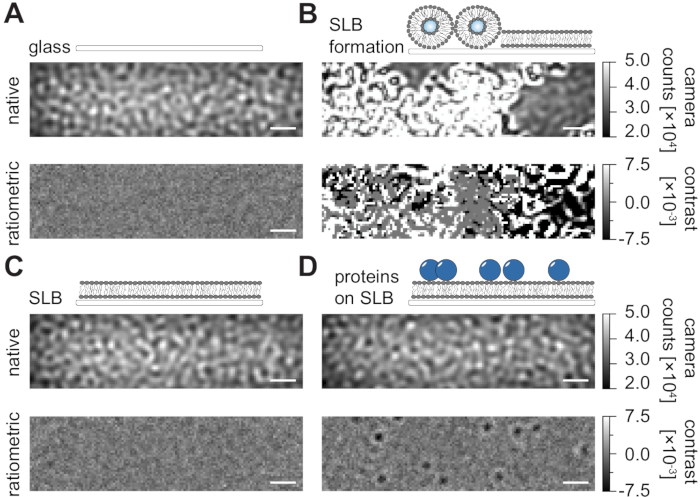
Figure 2: Native and ratiometric view of exemplary surfaces relevant for MSPT measurements. Representative images of the surface roughness of a glass cover slide (A), during the formation of a supported lipid bilayer (B), with an intact supported lipid bilayer (C) and of exemplary proteins reconstituted on an SLB (D). All four examples are displayed in the native mode, which can be accessed during the measurement itself, and as processed ratiometric images after median-based background removal. Scale bars represent 1 µm. For data analysis (see accompanied Jupyter notebook; step 9), the following parameters were used: median window size (window_length) = 1001. Please click here to view a larger version of this figure.
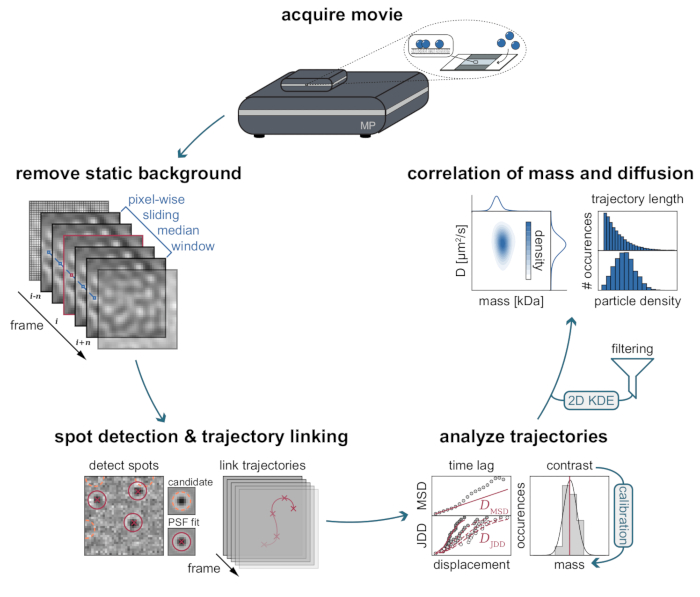
Figure 3: Step-by-step diagram of the stages required for MSPT data collection and analysis. After data acquisition for the sample of interest on the mass photometer, movies are processed to remove the static background through a pixel-wise sliding median approach. Thereafter, candidate particles are identified and fitted by a point spread function (PSF) prior to their linking into particle trajectories. To enable the determination of the diffusion coefficient for each particle, mean squared displacement (MSD) or jump distance distribution (JDD) analysis is employed. At this stage, contrast values can be transformed into molecular masses according to the contrast-mass-relation determined through the calibration strategy. As a final step, trajectories can be filtered based on their length or membrane particle density and visualized by two-dimensional kernel density estimation (2D-KDE). Please click here to view a larger version of this figure.
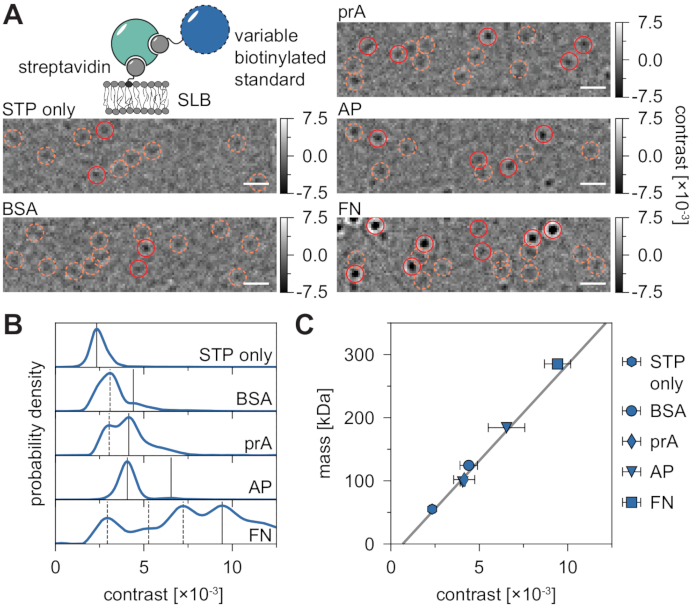
Figure 4: Calibration of the mass-to-contrast relation for MSPT measurements. (A) Representative ratiometric frames obtained for exemplary streptavidin-standard protein complexes diffusing on a supported lipid bilayer containing a small percentage of biotinylated lipids (DOPC:DOPG:Biotinyl Cap PE ratio of 70:29.99:0.01 mol%). As model molecular weight standards, monovalent streptavidin28 (STP only) or divalent streptavidin28 in complex with either biotinylated bovine serum albumin (BSA), biotinylated protein A (prA), biotinylated alkaline phosphatase (AP), or biotinylated fibronectin (FN) are shown. Candidate spots are highlighted in orange (dashed circles) and successful particle detections are highlighted in red (solid circles). The scale bars represent 1 µm. (B) Probability density distributions of contrast values obtained for the five model standard proteins. All displayed data represent pooled distributions of three independent experiments per condition: STP only n = 82,719; BSA n = 9,034; prA n = 22,204; AP n = 69,065, and FN n = 71,759 trajectories. As compared to particle numbers determined for membranes with proteins, the number of particles detected on an empty bilayer is negligible at moderate membrane densities (Supplementary Figure 1). Contrast peaks considered for mass calibration are marked through continuous lines while dashed ones represent oligomer states not considered. (C) Contrast-to-mass calibration curve derived from peak contrasts in panel D and the respective sequence masses of the complexes. Error bars display the standard error of the peak locations estimated by bootstrapping (100 resamples with 1,000 trajectories each). For data analysis (see Jupyter notebook; step 9), the following parameters were used: median window size (window_length) = 1,001 frames, detection threshold (thresh) = 0.00055, search range (dmax) = 4 pixels, memory (max_frames_to_vanish) = 0 frames, minimum trajectory length (minimum_trajectory_length) = 7 frames (STP only), 9 frames (BSA/FN), 15 frames (prA), 10 frames (AP). Please click here to view a larger version of this figure.
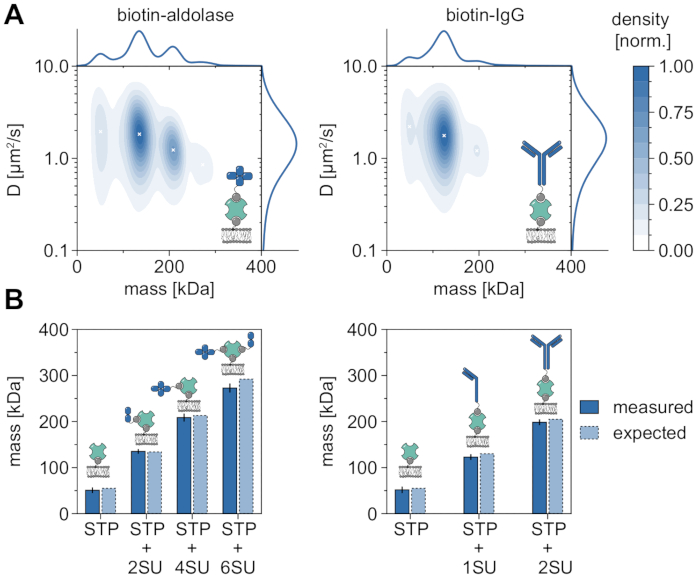
Figure 5: Deciphering oligomer states of membrane-associated proteins. (A) 2D kernel density estimations of both mass and diffusion coefficient of tetravalent streptavidin in complex with biotinylated aldolase (left panel) or with a biotin-modified goat anti-Rabbit IgG antibody (right panel). Reconstitution of both complexes has been performed on a supported lipid bilayer containing DOPC, DOPG, and Biotinyl Cap PE in a ratio of 70:29.99:0.01 mol%, respectively. In total, 116,787 trajectories of three independent replicates have been included for the streptavidin-aldolase complex (particle density of 0.1 µm-2) and 348,405 for the streptavidin-IgG composite (particle density of 0.1 µm-2). Only particles with a track length of at least five frames were included. Marginal probability distributions of both molecular mass (top) and diffusion coefficient (right) are presented. The black x in both panels mark the respective local maxima of the KDE. (B) Comparison of determined oligomer masses for the complex of tetravalent streptavidin with biotin-modified aldolase (left panel) or biotinylated IgG (right panel) with, according to the sequence masses, expected molecular weights. The abbreviation SU is introduced on behalf of the protein of interests' subunit. Error bars display the standard error of the peak locations estimated by bootstrapping (100 resamples with 1,000 trajectories each). For data analysis (see accompanied Jupyter notebook; step 9), the following parameters were used: median window size (window_length) = 1,001 frames, detection threshold (thresh) = 0.00055, search range (dmax) = 4 pixels, memory (max_frames_to_vanish) = 0 frames, minimum trajectory length (minimum_trajectory_length) = 5. Please click here to view a larger version of this figure.
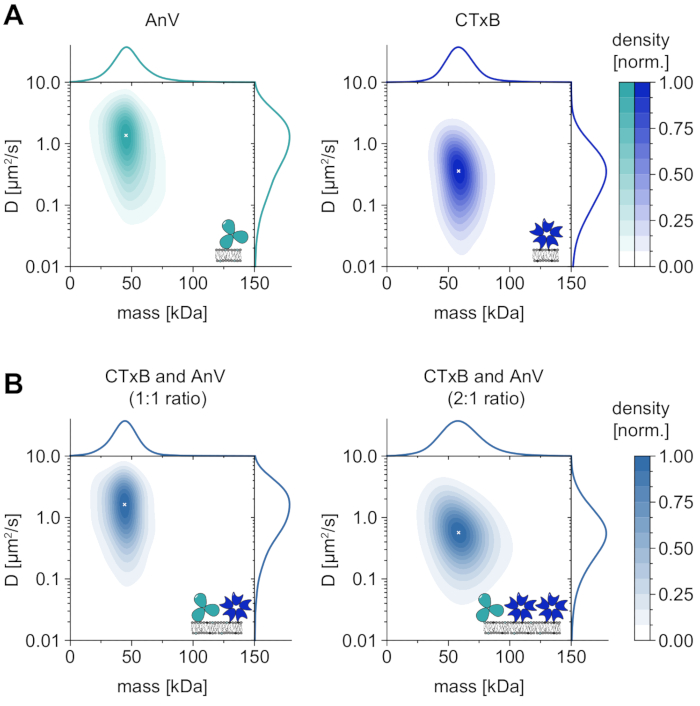
Figure 6: Dissolving the diffusive behavior of the native membrane-interacting proteins annexin V (AnV) and cholera toxin subunit B (CTxB). (A) 2D kernel density estimations of both mass and diffusion coefficient of annexin V (left panel) and cholera toxin subunit B (right panel). For AnV and CTxB membrane reconstitution, lipid compositions of 80:20 mol% DOPC to DOPS and 99.99:0.01 mol% DOPC to GM1 have been used, respectively. In total 206,819 trajectories of three independent replicates have been included for AnV (particle density of 0.1 µm-2) and 142,895 trajectories for CTxB (particle density of 0.2 µm-2). (B) 2D kernel density estimations of CTxB and AnV mixtures in a ratio of 1:1 (left panel) or 2:1 (right panel), respectively. Reconstitution of protein mixtures was performed on a supported lipid bilayer containing DOPC, DOPS, and GM1 lipids in a ratio of 80:19.99:0.01 mol%. In total 42,696 trajectories of three independent replicates have been included for the 1:1 mixture (particle density of 0.1 µm-2) and 264,561 trajectories for the 2:1 ratio (particle density of 0.3 µm-2). For both (A) and (B), only particles with a track length of at least five frames were included. Marginal probability distributions of both molecular mass (top) and diffusion coefficient (right) are presented. The white x in each panel marks the respective global maximum of the KDE. For data analysis (see accompanied Jupyter notebook; step 9), the following parameters were used: median window size (window_length) = 1,001 frames, detection threshold (thresh) = 0.00055, search range (dmax) = 4 pixels, memory (max_frames_to_vanish) = 0 frames, minimum trajectory length (minimum_trajectory_length) = 5. Please click here to view a larger version of this figure.
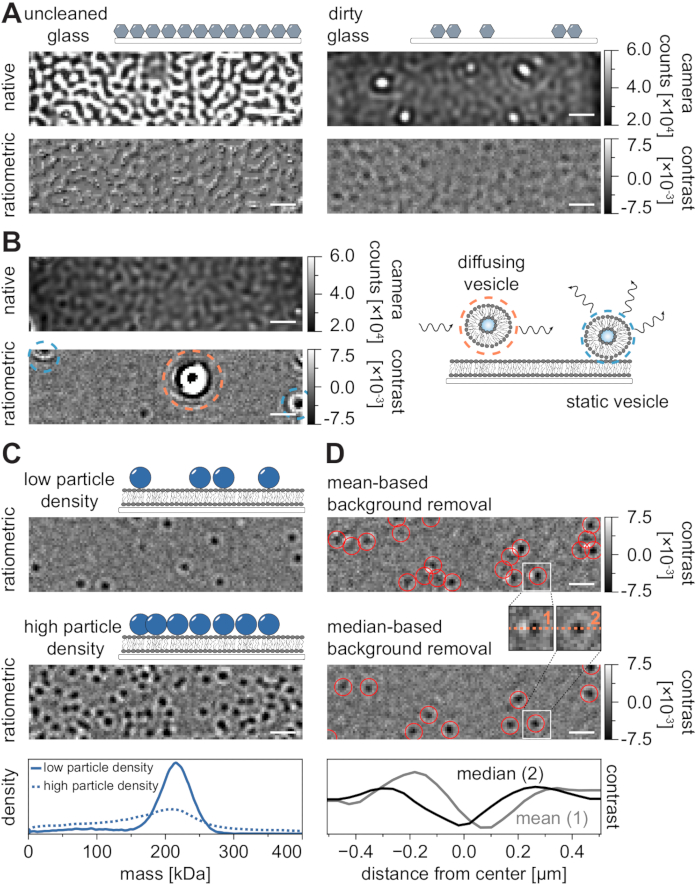
Figure 7: Potential complications in the course of MSPT measurements or during data analysis. (A) Representative images of the surface roughness displayed in both the native and the processed (median-based background removal) ratiometric view of an uncleaned cover glass slide. In both cases, bright spots constitute residual surface impurities that impede artifact-free measurements. (B) Exemplary images of residual vesicles in the field of view after insufficient membrane washing. Both static (highlighted in blue) and diffusing (highlighted in orange) vesicles will impair measurement quality either due to pulsating and wiggling or due to their directional movement, respectively. (C) As a single-particle technique, MSPT requires low particle densities (representative image, upper panel) to enable proper linking and mass determination of each particle. In the case of high membrane-particle densities (middle panel), particle fitting is impaired, which affects mass-determination (see lower panel). (D) Representative ratiometric images of particles diffusing on a membrane interface after either mean-based (upper panel) or median-based background removal. For diffusing particles, the mean-based background removal strategy produces distorted images of the particle's PSF as can be seen in the small insets between the upper and middle panel. In contrast, undistorted particle PSFs can be obtained through the median-based approach. Lower panel: Comparison of line profiles through the center of the PSF obtained after mean- or median-based background removal. For all native and ratiometric images displayed in this figure, the scale bars represent 1 µm. For data analysis (see accompanied Jupyter notebook; step 9), the following parameters were used: median window size (window_length) = 1,001 frames, detection threshold (thresh) = 0.00055, search range (dmax) = 4 pixels, memory (max_frames_to_vanish) = 0 frames, minimum trajectory length (minimum_trajectory_length) = 5. Please click here to view a larger version of this figure.
Supplementary Figure 1: Comparison of protein-free and occupied membranes. Representative images of an intact supported lipid bilayer before (A) and after (B) the addition of purified streptavidin (STP). Candidate spots that were successfully fit to the model PSF are encircled in red. (C) Contrast probability distributions of particles detected on an empty membrane (membrane background, gray) and on a bilayer with diffusing streptavidin particles (blue). Both probability distributions represent the pooled data of three independent experiments with identical movie acquisition and analysis parameters. For data analysis (see accompanied Jupyter notebook; step 9), the following parameters were used: median window size (window_length) = 1,001 frames, detection threshold (thresh) = 0.00055, search range (dmax) = 4 pixels, memory (max_frames_to_vanish) = 0 frames, minimum trajectory length (minimum_trajectory_length) = 7 frames. Please click here to download this File.
Supplementary Movie 1: Exemplary movie showing the rupture and fusion of vesicles into a homogeneous membrane recorded with the mass photometer. Image processing median window size (window_length) = 1,001 frames. Scale bar: 1 µm. Camera counts range: black = 16,892; white = 65,408. Please click here to download this Movie.
Supplementary Movie 2: Exemplary movies showing the diffusion of annexin V (top) and biotinylated aldolase (bottom) complexes on a bilayer as obtained from MSPT measurements. Image processing median window size (window_length) = 1,001 frames. Scale bar: 1 µm. Interferometric scattering contrast range: black = -0.0075; white = 0.0075. Please click here to download this Movie.
