The primary porcine RPE (pRPE) cells were cultured in DMEM/Basic media with 10% FBS, and cell morphology under light microscope was photographed at 2 days (Figure 2A), 6 days (Figure 2B) and 10 days (Figure 2C) after seeding. After 1 wk, a confluent monolayer of pigmented pRPE cells with cobblestone morphologies was observed.
To better characterize the primary pRPE cells, primary human RPE cells (hRPE) at Passage 3 (P3)18 and ARPE-19 cells were cultured in DMEM/Basic media with 1% FBS for another wk, and then the total mRNA and protein of the cultured cells, as well as porcine RPE/choroid tissues, were harvested. The expression levels of key characteristic genes19 and protein markers of mature RPE were evaluated by qRT-PCR and Western blot. Compared with ARPE-19 cells, pRPE cells retained significantly higher expression levels of genes functioning in native RPE secretion (Figure 3A(c)), phagocytosis (Figure 3A(i)), transportation (Figure 3A(a),(d)), tight junctions (Figure 3A(j)), barrier formation (Figure 3A(b)), as well as visual cycle (Figure 3A(e),B). However, the expression levels of Krt8 and Krt18, two cytoskeleton marker genes of the RPE, were significantly lower in primary pRPE cells in comparison with primary hRPE and ARPE-19 cells (Figure 3A(g),(h)). Moreover, itgav, which participates in phagocytosis, was lower in primary pRPE cells as well (Figure 3A(f)). Since the expression levels of these three genes in pRPE cells were similar with the porcine RPE/choroid tissue, this may indicate the gene expression differences between the species. Moreover, the expression level of Tyr, which is responsible for melanin production, was highly reduced in both primary pRPE and hRPE cells (Figure 3A(k)), which may explain the loss of pigmentation in long-term cultured cells. The dissimilarities in the qRT-PCR results observed in the data of human pRPE and porcine pRPE cells might be because of the longer storage and higher passage number (P3) of human pRPE, thus indicating a loss of RPE characters on sub-culturing. In addition, Western blot showed that RPE65 protein, which is a key enzyme in the visual cycle featuring RPE cells, was expressed in pRPE cells, while its expression level was significantly reduced in primary hRPE cells and ARPE-19 cells (Figure 3B,C).
To further characterize the polarity and barrier functions of RPE cells, the cultured confluent monolayers of pRPE and ARPE-19 cells were cultured in transwell inserts with DMEM/Basic, DMEM/F12, and MEM-Nic media for 1 wk (Figure 4). Under these culture conditions, Na+-K+-ATPase (Figure 4A–F) and ZO-1 (Figure 4G–L) fluorescent staining results revealed that pRPE cells had higher expression levels of both Na+-K+-ATPase and ZO-1 than ARPE-19 cells. The equal distribution of Na+-K+-ATPase at the apical and basal surface of pRPE cells indicated that a longer culture time was required to restore cell polarities in vitro.
The RPE forms tight junctions near its apical surface to tightly regulate the exchange of metabolites between the inner retina and choroids20. ZO-1 staining suggested that tight junction proteins were normally localized at the plasma membrane of primary pRPE cells with regular cobblestone morphologies, but not of ARPE-19 cells. Among the three culture medias, the best ZO-1 staining patterns were observed in cells cultured in DMEM/Basic, while DMEM/F12 failed to maintain the cobblestone morphology of pRPE cells. A higher TER value serves as an indicator of tight junction formation and better barrier functions of RPE cells21. To confirm the function of tight junctions, transepithelial resistance (TER) was measured. However, only slightly higher TER was detected in comparison with empty transwell inserts (Figure 4M,N).
In the retina, Bruch's membrane exists between the RPE and the choroid. The main components of Bruch's membrane are collagen type IV, proteoglycans, and laminin22. In order to simulate the supporting effect of Bruch's membrane on RPE, laminin was spread on the surface of the transwell membrane to facilitate the maturation of cultured RPE cells. Based on the results obtained from Figure 4, the confluent pRPE cells were cultured on transwell inserts in DMEM/Basic media with 1% FBS for 2 wks. Immunofluorescent staining results showed that RPE-specific proteins RPE65 were expressed in all pRPE cells (Figure 5A,B). Na+-K+-ATPase was distributed on the apical surface of pRPE cells, suggesting the re-establishment of cell polarities (Figure 5C,D). Both ZO-1 staining and TER results indicated that tight junctions were well formed in primary pRPE cells when they were cultured on laminin (Figure 5E,F,H). Figure 5G depicts cells stained with Hoechst dye.
Moreover, Western blot demonstrated that both pRPE and ARPE-19 cells produced growth factors, including pigment epithelium-derived factor (PEDF) and vascular endothelial growth factor (VEGF) (Figure 6A). After confluent monolayers of pRPE were cultured in transwell inserts with DMEM/Basic, DMEM/F12, and MEM-Nic media for 1 wk, culture media from the top and bottom chambers of the transwell inserts were collected, and the secreted VEGF was quantified by ELISA. When DMEM/Basic and MEM-Nic media were used for cell culture, pPRE cells secreted more VEGF than cells in DMEM/F12 media (Figure 6B). However, no differences could be detected in VEGF amounts between the top and bottom chambers of the transwell inserts in all tested conditions (Figure 6B). In addition, after primary pRPE cells were cultured on the inserts coated with laminin and in DMEM/Basic media for 2 wks, the media from the top and bottom chambers were collected. Western blot analysis showed higher levels of PEDF in the top chamber than the bottom chamber, while the protein levels of secreted VEGF were similar at both chambers (Figure 6C). These results further supported the re-establishment of cell polarity when primary pRPE cells were cultured for another 2 wks after they reached confluency on laminin.

Figure 1: Basic steps of pRPE cell isolation. (A) Wash porcine eyeballs with 1x PBS in 50 mL sterile centrifuge tubes. (B,C) Prepare RPE-choroid-sclera complexes for enzymatic digestion. Please click here to view a larger version of this figure.
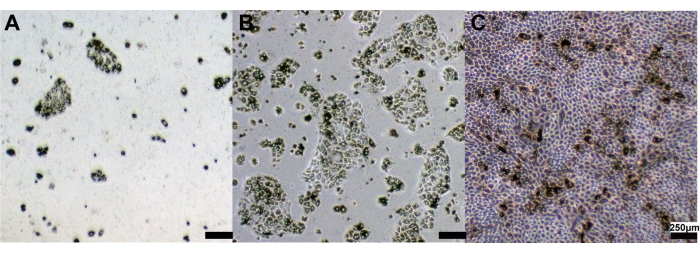
Figure 2: Primary pRPE cell morphologies after cell culture. Representative images of primary pRPE cells at (A) Day 2, (B) Day 6, and (C) Day 10. Scale bar 250 µm. Please click here to view a larger version of this figure.
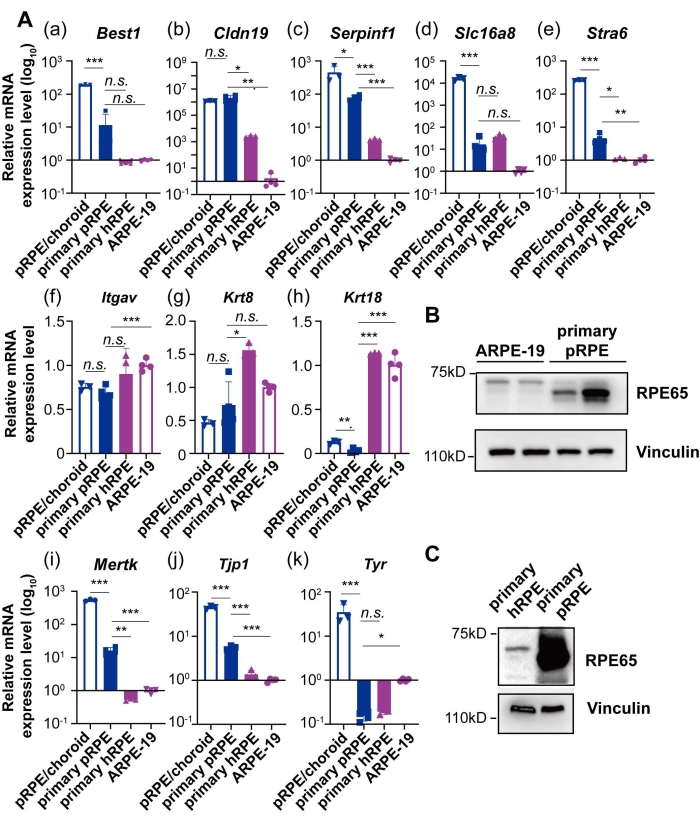
Figure 3: Expression levels of key signature genes and proteins in cultured RPE cells. (A) qRT-PCR analysis of mRNA levels of signature genes in primary pRPE cells, primary hRPE cells, ARPE-19 cells, and pRPE/choroid tissue. Gapdh was used as the house keeping gene for qRT-PCR. Four biological replicates were used for primary pRPE cells and ARPE-19 cells, while three biological replicates were used for primary hRPE cells and pRPE/choroid tissue. The gene expression levels in ARPE-19 cells were set as controls. (B) Western blot analysis of RPE65 proteins in ARPE-19 and pRPE cells. Vinculin was used as a loading control. (C) Western blot analysis of RPE65 proteins in primary hRPE cells and pRPE/choroid tissue. Vinculin was used as a loading control. For comparison, student t-test was used, *p < 0.05, **p < 0.01, ***p < 0.001. Please click here to view a larger version of this figure.
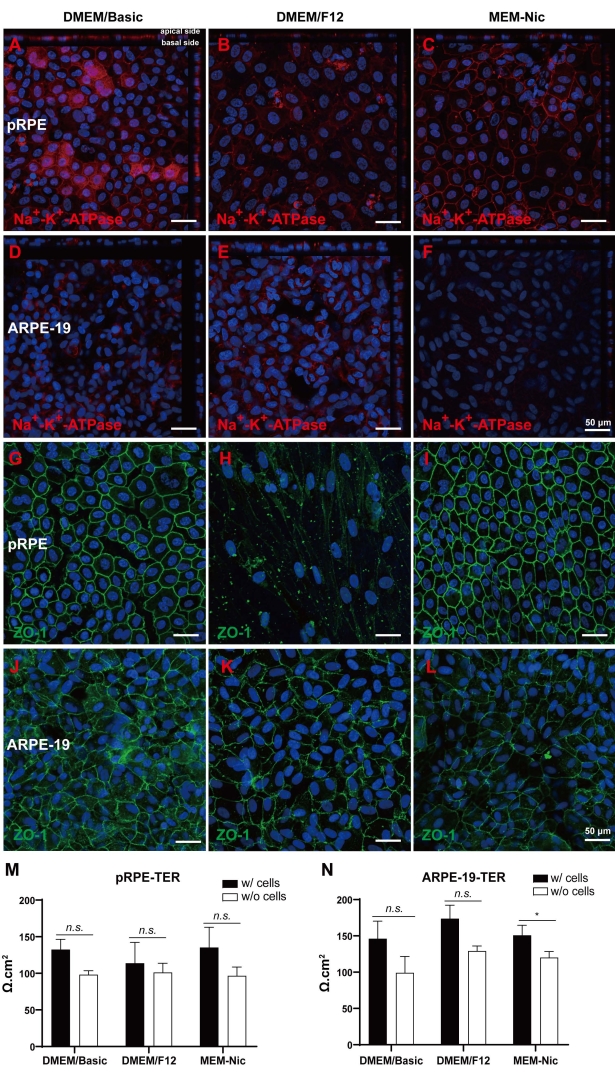
Figure 4: One-wk culture of primary pRPE and ARPE-19 cells in DMEM/Basic, DMEM/F12, and MEM-Nic media with 1% FBS. (A) Na+-K+-ATPase fluorescent staining (Red) of primary pRPE cells in DMEM/Basic, (B) DMEM/F12, and (C) MEM-Nic media. (D) Na+-K+-ATPase fluorescent staining (Red) of ARPE-19 cells in DMEM/Basic media, (E) DMEM/F12 media, and (F) MEM-Nic media. (G) ZO-1 fluorescent staining (Green) of primary pRPE cells in DMEM/Basic media, (H) DMEM/F12 media, and (I) MEM-Nic media. (J) ZO-1 fluorescent staining (Green) in ARPE-19 cells in DMEM/Basic media, (K) DMEM/F12 media, and (L) MEM-Nic media. TER measurements after primary pRPE (M) and ARPE-19 (N) cells were cultured in DMEM/Basic, DMEM/F12, and MEM-Nic media for 1 wk. For each type of cells, data were obtained from at least three different wells. For comparison, student t-test was used, *p < 0.05, **p < 0.01, ***p < 0.001. Scale bar 50 µm. Please click here to view a larger version of this figure.
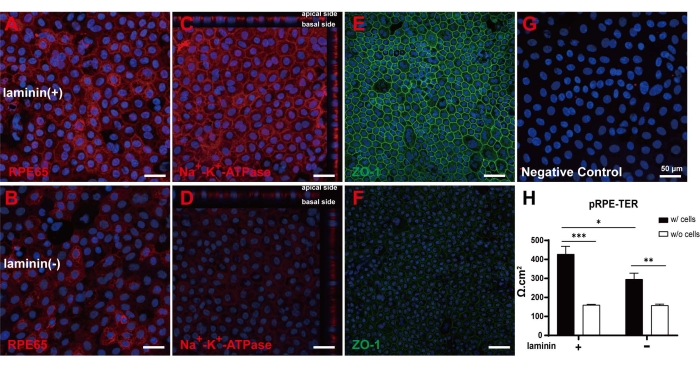
Figure 5: Two-wk culture of pRPE cells in DMEM/Basic media with 1% FBS. (A–F) Cells cultured with or without laminin were stained with RPE65 (A,B, Red), Na+-K+-ATPase (C,D, Red), ZO-1 (E,F, Green), and (G) Hoechst (Blue) . (H) TER measurements in cell sheets cultured with or without laminin. For different treatments, data were obtained from four different wells. For comparison, student t-test was used, *p < 0.05, **p < 0.01, ***p < 0.001. Scale bar = 50 µm. Please click here to view a larger version of this figure.
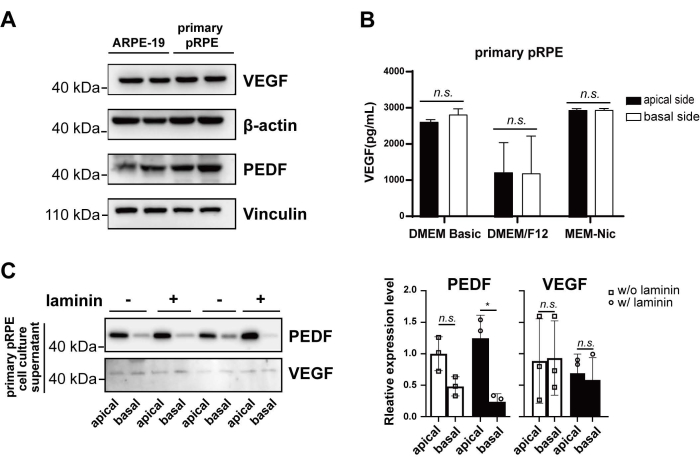
Figure 6: Secretory functions of cultured primary pRPE and ARPE-19 cells. (A) Western blot analysis of VEGF and PEDF in primary pRPE and ARPE-19 cells. (B) Secreted VEGF in the top and bottom chamber of transwell inserts with primary pRPE cells when confluent cells were cultured in DMEM/Basic, DMEM/F12, and MEM-Nic media for 1 wk. (C) Secreted PEDF and VEGF at the top and bottom chamber of transwell inserts with primary pRPE in DMEM/Basic media for 2 wks. For different treatments, data were obtained from three different wells. For comparison, student t-test was used, *p < 0.05, **p < 0.01, ***p < 0.001. Please click here to view a larger version of this figure.
| Homo sapiens | Sus scrofa | |
| Best1 | F: GAAGGCAAGGACGAGCAAG | F: GGACACCTGTATGCCTACGA |
| R: TCCAACTGCTTGTGTTCTGC | R: GGAACGTGAAGAGGGGTACA | |
| Cldn19 | F: GGTGACCCAGGAGTTCTTCA | F: TCGTGACCCAGGAGTTCTTC |
| R: CTGTTGGGTCTCTCTGGCTC | R: GCTGCTGTTGGATCGCTC | |
| Itgav | F: CGCAGTCCCATCTCAAATCC | F: GCTTTCTTCAGGACGGAACA |
| R: GGCCCTGTATAAGATAGCTCGA | R: GAAATGAGCTGACCTTGCCA | |
| Krt8 | F: GGAGCAGATCAAGACCCTCA | F: CCCAGGAGAAGGAGCAGATC |
| R: GCCGCCTAAGGTTGTTGATG | R: ATGTTGTCGATGTTGCTCCG | |
| Krt18 | F: CTTGGAGAAGAAGGGACCCC | F: GCTGATAATCGGAGGCTGGA |
| R: GGCCAGCTCTGTCTCATACT | R: GAAGTCATCAGCAGCGAGAC | |
| Mertk | F: GTGTGCAGCGTTCAGACAAT | F: GCGGCTATTTCTTGGTGGAA |
| R: AAAATGTTGACGGGCTCAGG | R: ACGTAGATGGGGTCAGACAC | |
| Serpinf1 | F: CAGATGAAAGGGAAGCTCGC | F: AAGACGTCGCTGGAGGATTT |
| R: TTAGGGTCCGACATCATGGG | R: GGTCACTTTCAGAGGCAGGA | |
| Slc16a8 | F: GGGTGTCCTCCATCATGCTA | F: CAGTTCGAGGTGCTCATGG |
| R: GTCAGGTAGAGCTCCAGGAG | R: GACAGCCATGAAGACACCAG | |
| Stra6 | F: CTTTGCAGGAAGAAGCTGGG | F: CTAGCCGTGTTGTCGATCCT |
| R: TAAATGGCCGTCCCTGTCAG | R: GACCATGAAGACAGCAGCAG | |
| Tjp1 | F: ACAGGAAAATGACCGAGTTGC | F: AAGACTTGTCAGCTCAGCCA |
| R: TGGTTCAGGATCAGGACGAC | R: CCAGCATCTCGAGGTTCACT | |
| Tyr | F: ACTCAGCCCAGCATCATTCT | F: ATCTACTCAGCCCAGCATCC |
| R: ACATCAGCTGAAGAGGGGAG | R: GAGCCTTGGAGTCCTGGATT | |
| Gapdh | F: CAGCCTCAAGATCATCAGCA | F: CATCCTGGGCTACACTGAGG |
| R: ATGATGTTCTGGAGAGCCCC | R: GGGGCTCTTACTCCTTGGAG |
Table 1: Primers for qRT-PCR.
