For all experimental procedures, obtain approval from the Institutional Animal Care and Use Committee (IACUC) and adhere to NIH Guidelines for the care and use of laboratory animals. In the United States of America, GPs are additionally subject to United States Department of Agriculture (USDA) regulations. All the procedures in this protocol were approved by the University of Pittsburgh IACUC and adhered to NIH Guidelines for the care and use of laboratory animals. For this experiment, three male wildtype, pigmented GPs between 4 and 10 months of age, with ~600-1,000 g weight were used.
1. Surgical procedure
- Perform all pupillometry experiments in awake, head-fixed, and passively listening pigmented GPs. Verify normal hearing in experimental subjects using click and pure-tone auditory brainstem response (ABR) recordings58.
NOTE: Although pupillometry data acquisition is by itself non-invasive, an invasive head post implant surgery is used in this protocol for immobilizing the animal's head during the procedure. Alternatives are presented in the Discussion section. - First, implant all the experimental animals with a stainless-steel head post for head fixation under isoflurane anesthesia. Use aseptic surgical techniques to anchor the head post to the skull employing a combination of bone screws and dental acrylic58.
- Provide the animals with post-surgical care, including the administration of systemic and topical analgesics. After a 2-week recovery period, gradually acclimatize animals to the experimental setup.
NOTE: The surgical procedure is based on previously published methods in GPs58 as well as other species59,60, and is not the focus of this protocol.
2. Animal acclimation to the experimental setup
NOTE: Experiments typically take place in a sound-attenuated chamber or booth (see Table of Materials). The time required to familiarize an animal to the setup varies from subject to subject. Typical acclimation times are noted below. A well-acclimated animal will tolerate head-fixation with minimal body motion, and result in better pupil diameter measurements.
- After a 2-week recovery period, first familiarize the animals to handling and transport (2-3 days). This acclimation is essential to reduce stress and anxiety. To familiarize the animal to handling, place the animal in its transport container for increasing amounts of time (10-30 min), and handle the animal for increasing amounts of time (10-30 min).
- Next, acclimate the animal to the experimental setup (2-3 days) by placing the animal in an enclosure for 10-45 min (Figure 1A). The enclosure must allow for small postural shifts for the animal's comfort during the experiment. Allow for small postural shifts for the animal's comfort during the experiment. However, pupil dilation is known to precede motion49. Therefore, measure the motion of the animal and account for this motion in the data analysis (Figure 1C).
- As part of this acclimation, manually handle the implanted head post, as if the animal is going to be head fixed. Hold the head post for increasing durations (10-60 s).
- After manual acclimation and depending on animal behavior, try to head fix the animal to a rigid frame using the implant holder.
- Slowly increase the head fixation duration (10-45 min) until the animal is calm and relatively still while being head-fixed (2-3 days).
- Accustom the animal to the presence of the camera, IR light source, and white light source (1-2 days). Turn on the white light, gradually increasing the duration (10 min to 30 min).
- Accustom the animal to acoustic stimulation by playing a variety of sounds (e.g., pure tones, clicks, vocalizations) at different sound levels (1-2 days, concurrent with step 2.6). To minimize habituation to experimental stimuli, use sounds different from those planned for the pupillometry experiments in this step.
3. Calibration of pupil camera
NOTE: The camera used for pupillometry outputs a video via USB to the pupillometry software suite. From this video, the pupil diameter is extracted using an ellipse fit and user-adjustable threshold value by the pupillometry software suite (see Table of Materials). The software then interfaces with a digital-to-analog card. The card outputs an analog voltage value that is proportional to the pupil diameter. Calibration is needed to convert this voltage value back to pupil diameter in units of length.
- Place a sheet of paper containing images of black discs of known diameter at the same location where the GP's eye will be located during pupillometry. For GPs, PD is in the 4 mm range. Therefore, perform calibration using 3 mm, 4 mm, and 5 mm discs.
- Place the pupillometry camera (see Table of Materials) at the same distance (25 cm) at which the experiments will be performed. Adjust the camera aperture and focus until a sharply focused image of a disc of known diameter is obtained.
- In the pupillometry acquisition software (see Table of Materials), adjust the threshold so that the outline of the ellipse fit closely matches the imaged disc, and note down the analog output voltage value and scaling.
- Repeat this procedure for the 3 mm, 4 mm, and 5 mm discs. Then, tabulate the actual diameter values (in mm) corresponding to the analog output voltage values.
4. Pupillometry data acquisition
- Perform all the experiments in a sound-attenuated booth or chamber, with the inner walls covered with anechoic foam.
- For free-field stimulus delivery, mount a calibrated loudspeaker onto the sound attenuated chamber wall, at an equal height to the position where the animal will be placed.
NOTE: The choice of loudspeaker depends on the species being studied and the stimuli planned. For GP vocalizations, use a full-range driver speaker that has a relatively flat (±3 dB) frequency response in the vocalization frequency range of 0.5-3 kHz (Figure 1A). - Place the animal in the enclosure ensuring that large body movements are not possible (Figure 1A).Fix the head of the animal to the rigid frame as described in step 2 (Figure 1A).
- Place a piezoelectric sensor beneath the enclosure in order to detect and record animal movements (Figure 1A).
- To set up the air puff, use a holder attached to the tabletop to place a pipette tip at ~15 cm in front of the animal's snout. Connect a silicon tube (~3 mm diameter) to the pipette tip and connect the tube to a regulated air cylinder.
- Keep the cylinder air pressure between 20 and 25 psi. Pass the tube through a pinch valve to control the timing and duration of the air puff using a computer-controlled relay.
- Illuminate the eye with an infrared LED array placed at ~10 cm distance. Use white LED lighting at an intensity of ~2,000 cd/m2 to illuminate the imaged eye and bring the baseline PD to ~3.5 mm. Maintain constant illumination conditions in the experimental chamber across experimental sessions.
NOTE: In normal laboratory lighting (~500 cd/m2), the GP pupil is quite dilated, and does not allow for the observation of further stimulus-linked dilation. By using additional illumination, the pupil is brought to a baseline diameter of ~3.5 mm, allowing for a sufficient dynamic range to observe stimulus-linked dilation. This also ensures consistent baselines across sessions and subjects. - Open the pupil acquisition software and acquire the video (at 90 fps) of the pupil using a camera with a 16 mm lens (spatial resolution of 0.15° visual angle) and infrared (IR) filter placed at a 25 cm distance from the imaged eye. Ensure that the eye is centered in the imaged area.
- Regulate the aperture and focus of the camera, as well as the IR level until the outline of the imaged pupil is in sharp focus.
- In the pupil acquisition software, define the area of interest containing the pupil by selecting a rectangular area with the mouse.
- Use the pupil acquisition software controls panel to adjust the brightness and contrast of the acquired video. Set the scan density to 5 and adjust the threshold such that the ellipse fit closely matches the outline of the pupil in the video.
- Using the neural interface processor software, acquire and save the analog signal from the PD trace, the voltage trace from the piezoelectric sensor recording motion, the stimulus delivery times, and the air puff delivery times.
5. Call-in-noise detection and categorical discrimination using a modified oddball paradigm
NOTE: The stimuli for pupillometry experiments consisted of GP vocalizations that were recorded in an animal colony58. The vocalization samples can be found in the following repository: https://github.com/vatsunlab/CaviaVOX. In particular, wheek and whine calls were used to elicit the pupil responses shown in the representative results. From each category, choose vocalizations whose lengths are approximately equal. To account for differences in the recording amplitude and temporal envelopes of the vocalizations, normalize the vocalizations by their root mean square (r.m.s.) amplitudes, if needed.
- Present the auditory stimuli using MATLAB at an appropriate sampling rate. For GPs, which are low-frequency-hearing animals, a sampling rate of 100 kHz is sufficient.
- Select eight different exemplars of GP vocalizations of similar lengths from two different categories of vocalizations (e.g., wheek calls and whine calls). One category (eight exemplars) will serve as standard stimuli, and the other category (eight exemplars) will serve as the oddball or deviant stimuli (Figure 2A).
- To generate 1 s long standard and deviant stimuli embedded in noise at different signal-to-noise-ratio (SNR) levels, add white noise of equal length to the calls (gated noise). The range of SNRs sampled in this experiment is between -24 dB SNR and +40 dB SNR.
- Using a block design, in each experimental session (~12 min duration), acquire data corresponding to a single SNR level. In each session, use eight exemplars of one vocalization category at a particular SNR as standard stimuli, and eight exemplars of the other vocalization category at that same SNR level as deviant stimuli.
NOTE: A typical experimental block lasts ~12 minutes. Depending on the animal's behavior and habituation of pupil responses, it may be possible to acquire data for 3-4 blocks each day (~45 – 60 mins). Throughout this duration, monitor the animal closely via the pupil video, the motion trace, as well as directly between blocks. - For each session, prepare a pseudorandom stimulus presentation sequence that contains standard stimuli >90% of the time. Ensure that between deviant stimuli, there are at least 20 trials with standard stimuli (Figure 2B).
NOTE: Depending on the experiment, the ordering of deviant stimuli within the stimulus presentation sequence can adopt a Latin square design to ensure that each unique deviant stimulus occupies a unique sequential position in every session. Averaging over all sessions can thus minimize the effect of the deviant stimulus position within the overall stimulus sequence. - Use a fixed stimulus intensity (for example, 85 dB SPL) for all stimulus presentation.
NOTE: Use an appropriate digital-to-analog converter to generate an audio signal, attenuate it to the desired sound level using a programmable attenuator, power-amplify the signal and deliver the signal using a calibrated loudspeaker (for example, hardware, see Table of Materials). - Present the stimuli with high temporal regularity (1 s stimulus followed by 3 s of silence as shown in the representative results).
NOTE: The pupil dilation responses are slow, typically peaking about 1 s after stimulus onset and taking about 5 s to return to baseline49. The stimulus presentation rate must be low enough to account for these slow timescales. Temporal regularity is important because it is possible that interrupting the timing pattern could itself act as a deviant stimulus. - To maintain the animal's engagement with the stimuli and to minimize habituation, optionally deliver a brief air puff (100 ms) after the deviant stimulus. Ensure that the onset of the air puff is sufficiently separated from the stimulus duration (2.5 s from stimulus onset) so that stimulus-evoked pupil dilation responses reach a peak before air puff induced blink artifacts.
NOTE: In the classical oddball paradigm, no positive or negative reinforcements are used. Since an air puff is used here as a mildly aversive reinforcement to maintain the animal's engagement with the auditory stimuli, the paradigm is referred to as a modified oddball paradigm.
6. Analysis and statistics
NOTE: All the analyses were performed using custom code written in MATLAB (available at https://github.com/vatsunlab/GP_Pupil). Two main analysis methods are described, which address the reliability and the time course of pupil responses, respectively. The choice of one or both the methods will be dictated by experimental design.
- Motion detection and trial exclusion
- Using the code pupil_avg_JOVE.m, perform motion detection and trial exclusion for every session. To do so, run the code and select the data file from a single session in the popup dialog.
- Linearly detrend the PD trace and convert the units from voltage to micrometers using the calibration table derived earlier (see step 3). Also, linearly detrend the motion trace over the entire recording session (~12 min).
- Inspect the session data by plotting the pupil trace (Figure 1B – top row) and the linearly detrended motion trace (Figure 1B– bottom row) over the session duration (~12 min), overlaid on trial markers.
- Measure the standard deviation (SD) of the motion trace. Obtain the times of motion trace peaks using the findpeaks function in MATLAB. Consider the peaks that crossed a threshold of 5 SDs and that are separate from other peaks by at least 1 s as a motion event49 (Figure 1B – bottom).
- Discard any trials (both standard and deviant) of pupil dilation that occurs within 7 s of a motion event. If more than half the number of deviant trials is discarded due to motion-related pupil dilation, discard the entire session and repeat it.
- Data pre-processing and visualization
- Use the code pupil_avg_JOVE.m, to remove eye blink artifacts, pre-process the data, and obtain the average pupil dilation to each stimulus across sessions. To do so, run the code and select all the data files to be analyzed in the popup dialog.
- Detect eye blinks (PD changes exceeding 400 µm/ms) and remove them by linearly interpolating the PD trace in a 200 ms time window centered at the detected blink time. Discard the session data if more than half the number of deviant trials contain an eye blink between stimulus onset and air puff onset.
- Downsample PD data from the acquisition sampling rate of 1,000 Hz to 10 Hz.
- Extract PD traces in a window beginning 1 s before the stimulus onset and lasting 5 s after stimulus offset. Compute the average baseline PD for each stimulus in a 500 ms window just prior to the onset of the stimulus. Subtract the baseline PD from these traces to obtain the stimulus-evoked change in PD.
- Average the stimulus-evoked PD changes for each stimulus condition across sessions within each animal, and then across animals to generate the mean pupil dilation response to each stimulus condition (for example, Figure 3A).
- Growth curve analysis (GCA) for quantifying the time course of PD changes
NOTE: This analysis method determines the magnitude and time course of pupil dilation responses and has been used in pupillometric studies of human subjects27,36,40 as well as in guinea pigs49.- Vertically concatenate all the outputs from pupil_avg_JOVE.m for all the sessions, animals, SNRs and attenuations to construct a matrix containing the following columns: animalID, SNR, sound level, and Pupil (1-50) diameter values. Using the code pupil_LME_JOVE.m, perform the growth curve analysis (GCA)27,36,40,49.
- Fit linear mixed-effect models with subject-level intercepts as random effects, and orthogonal time polynomials of up to order two as fixed effects, with each deviant SNR treated as a separate group, to the rising phase of the pupil diameter trace (0.1 to 2.1 s following stimulus onset).
- Model the rising phase of the pupil trace using the following formula36,49:
Pupildilation = (Intercept + Condition) + time1 * (βtime1 + βtime1: Condition) + time2* (βtime2+ βtime2: Condition) + r(subjectlevelintercept)
Where, time1 and time2 correspond to orthogonal linear and quadratic time polynomials, and βs correspond to weights. - Estimate mean weights (βs) and their standard errors using the fitlme function in MATLAB. Estimate the statistical significance of the weights using the coeftest function.
- For each SNR, plot the weights corresponding to the intercept, linear, and quadratic terms to visualize the results (Figure 3B, C).
- Analysis of trials showing statistically significant pupil dilations
NOTE: This analysis method determines the fraction of deviant trials on which a statistically significant pupil dilation response is observed and corresponds to the reliability of pupil dilation responses.- Choose an appropriate analysis window (0.5-1 s) centered around the peak of the pupil response (usually ~1.5 s after stimulus onset). Compute the mean PD in this analysis window for all standard and deviant trials.
- Determine whether the mean PD for each of the deviant trials is greater than 2.33 standard errors of the pooled distribution of mean PD values for standard trials. Count the deviant trials that exceed this threshold as trials showing a significant pupil dilation.
- Divide the number of deviant trials showing a significant pupil dilation by the total number of deviant trials (for each condition) to quantify the fraction of trials that show statistically significant increases in PD compared to standard stimulus trials.
- Put all the session-wise percentage of trials with significant pupil changes into each cell of a cell-array, where the cells are arranged from lower to higher SNR. Using the code pupil_threshold_estimate_JOVE.m, estimate the call-in-noise-categorization threshold.
- Plot the fraction of trials that show a statistically significant increase in PD as a function of SNR (Figure 3D). To these data, use the fitnlm MATLAB function (in the statistics toolbox) to fit psychometric functions of the form61:
Ψ(x; α, β, λ) = (1 -λ) * F(x; α, β)
Where, F is the Weibull function, defined as
F(x; α, β) =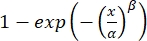 , α is the shift parameter, β is the slope parameter, and λ is the lapse rate.
, α is the shift parameter, β is the slope parameter, and λ is the lapse rate.
Pupillometry was performed in three male pigmented GPs, weighing ~600-1,000 g over the course of the experiments. As described in this protocol, to estimate call-in-noise categorization thresholds, an oddball paradigm was used for stimulus presentation. In the oddball paradigm, calls belonging to one category (whines) embedded in white noise at a given SNR were employed as standard stimuli (Figure 2A), and calls from another category (wheeks) embedded in white noise at the same SNR (Figure 2A) as deviant stimuli. Standard and deviant stimuli were randomly chosen, with resampling, from eight exemplars of each category. In each experimental session, stimuli were presented with high temporal regularity (Figure 2B), with at least 20 presentations of standard stimuli between deviant stimuli. Data were acquired corresponding to a particular SNR level in each experimental session. Across sessions, a broad range of clean and noisy SNRs were sampled (-24, -18, -12, -6, -3, 0, 3, 6, 12, 40 dB SNR).
The PD changes to the standard stimuli did not differ significantly from the baseline (blue line in Figure 3A). The deviant stimuli evoked robust and significantly larger PD changes than those elicited by the standard stimuli (gray lines in Figure 3A), reflecting call category discrimination. The response magnitude and the percentage of trials with statistically significant pupil responses were highest at the cleanest SNR and decreased gradually with decreasing SNR (Figure 3A,B). Using GCA, pupil responses to deviant stimuli were found to be statistically significant at SNRs above -18 dB (Figure 3C), which was taken to be the call-in-noise categorization threshold (green line in Figure 3A). The percentage of significant trials at each tested SNR level was well-fit by a psychometric function (Figure 3D). The SNR level necessary to reach the half-maximum of the psychometric curve was about -20 dB SNR (Figure 3D). Anecdotally, for this case, the reliability-based and time course-based metrics yielded similar values of call-in-noise categorization thresholds.
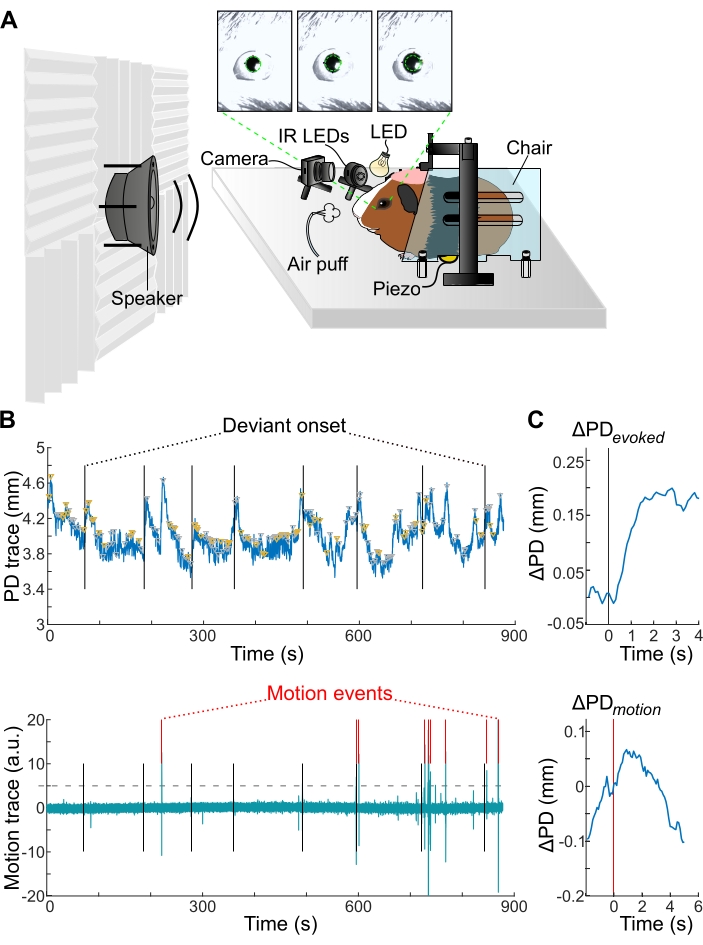
Figure 1: Pupillometry setup, and stimulus-evoked and motion-related PD changes. (A) The pupillometry setup with video frame images of sound evoked pupil dilation (top). The baseline PD is shown by dashed green circles. (B) An exemplar PD trace (top) and exemplar motion trace (bottom) from a single experimental session. Vertical black lines correspond to onset time deviant stimulus presentations. Red ticks correspond to automatically detected motion events. Gray horizontal dashed line corresponds to 5 SD threshold. (C) The PD changes (ΔPD) evoked by deviant stimulus (top) and related to motion events (bottom) from one experimental session. Stimulus onset is shown by vertical black line; the detection of motion event is shown by vertical red line. Note that pupil dilation onset precedes the onset of motion. Please click here to view a larger version of this figure.
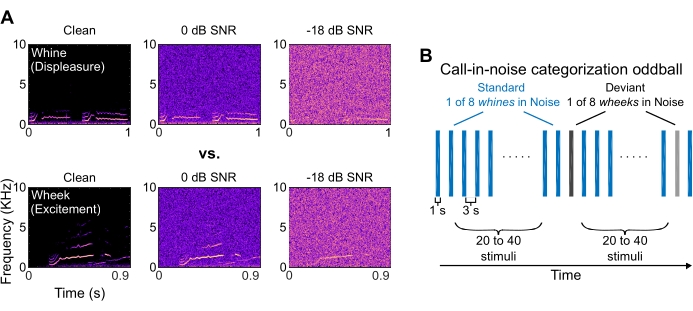
Figure 2: Call spectrograms and call-in-noise categorization paradigm structure. (A) Representative spectrograms of a guinea pig whine and wheek, in clean conditions and at 0- and -18-dB SNR, respectively. Noisy calls were obtained by adding white noise. (B) Structure of the oddball paradigm used to estimate call-in-noise categorization thresholds. Whine calls were randomly chosen from eight exemplars and used as standard stimuli. Wheek calls were randomly chosen from eight exemplars and used as deviants. In each experimental session, the noise was added at a different SNR level (-24, -18, -12, -6, -3, 0, 3, 6, 12 dB SNR). The calls are 1 s long and the time between stimuli is 3 s. Please click here to view a larger version of this figure.
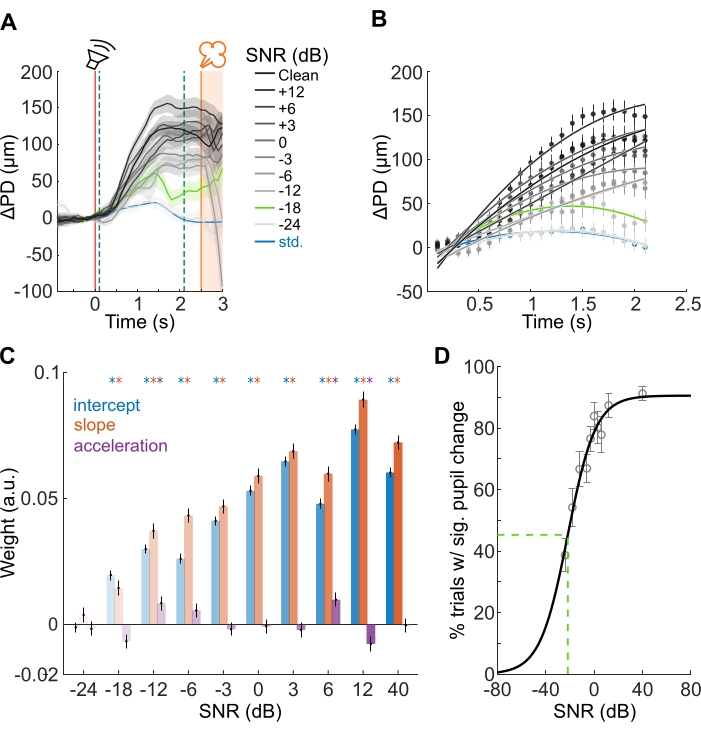
Figure 3: Pupillometry estimates of call-in-noise detections and categorization thresholds. (A) Average pupil responses from three animals. Mean pupil responses to standard whine stimuli are represented by blue line, and shading corresponds to ±1 standard error of mean (s.e.m.). Gray lines and shading correspond to mean and ±1 s.e.m. of pupil responses evoked by deviant wheek stimuli. Gray shading intensity corresponds to SNR. Green line and shading correspond to average pupil trace at threshold SNR (about -18 dB SNR). Red vertical line corresponds to stimulus onset; orange vertical line corresponds to air puff onset; teal dashed lines correspond to GCA window (PD changes rising phase). (B) GCA fit to the rising phase of PD changes. Dots are mean pupil diameter in 100 ms time bins, whiskers correspond to ±1 s.e.m. Solid lines correspond to mixed-effects model fits. Line colors as in A. (C) GCA weight estimates. Weights of the intercept is in blue, slope is in red, and acceleration is in purple. Whiskers correspond to ±1 s.e.m. Asterisks show statistically significant regression weights (linear hypothesis test on linear regression model coefficients). (D) Psychometric function fit to the percent of trials with significant PD changes elicited by the deviant stimulus as a function of SNR. Whiskers correspond to ±1 s.e.m. Note that 50% of the maximum is reached at about -20 dB SNR (green dashed line). Please click here to view a larger version of this figure.
