Effective Techniques for the Feeding and Ex Situ Culture of a Brooding Scleractinian Coral, Pocillopora acuta
Summary
Climate change is impacting coral reef ecosystems globally. Corals sourced from ex situ aquaculture systems can help support restoration and research efforts. Herein, feeding and coral culture techniques that can be used to promote the long-term maintenance of brooding scleractinian corals ex situ are outlined.
Abstract
Climate change is affecting the survival, growth, and recruitment of corals globally, with large-scale shifts in abundance and community composition expected in reef ecosystems over the next several decades. Recognition of this reef degradation has prompted a range of novel research- and restoration-based active interventions. Ex situ aquaculture can play a supporting role through the establishment of robust coral culture protocols (e.g., to improve health and reproduction in long-term experiments) and through the provision of a consistent broodstock supply (e.g., for use in restoration projects). Here, simple techniques for the feeding and ex situ culture of brooding scleractinian corals are outlined using the common and well-studied coral, Pocillopora acuta, as an example. To demonstrate this approach, coral colonies were exposed to different temperatures (24 °C vs. 28 °C) and feeding treatments (fed vs. unfed) and the reproductive output and timing, as well as the feasibility of feeding Artemia nauplii to corals at both temperatures, was compared. Reproductive output showed high variation across colonies, with differing trends observed between the temperature treatments; at 24 °C, fed colonies produced more larvae than unfed colonies, but the opposite was found in colonies cultured at 28 °C. All colonies reproduced before the full moon, and differences in reproductive timing were only found between unfed colonies in the 28 °C treatment and fed colonies in the 24 °C treatment (mean lunar day of reproduction ± standard deviation: 6.5 ± 2.5 and 11.1 ± 2.6, respectively). The coral colonies fed efficiently on Artemia nauplii at both treatment temperatures. These proposed feeding and culture techniques focus on the reduction of coral stress and the promotion of reproductive longevity in a cost-effective and customizable manner, with versatile applicability in both flow-through and recirculating aquaculture systems.
Introduction
Many coral reef ecosystems globally are being lost and degraded as a result of high-temperature stress driven by climate change1,2. Coral bleaching (i.e., the breakdown of the coral-algal symbiosis3) was considered relatively rare in the past4 but is now occurring more frequently5, with annual bleaching expected to occur in many regions by mid to late century6,7. This shortening of the interim period between bleaching events can limit the capacity for reef resilience8. The direct impacts of high-temperature stress on coral colonies (e.g., tissue damage9; energy depletion10) are intrinsically linked to indirect impacts at the reef-scale level, of which a reduction in reproductive/recruitment capacity is of particular concern11. This has spurred a range of applied research exploring, for example, the active in situ enhancement of recruitment (e.g., reef seeding12), new technologies for scaling-up coral restoration13, and the simulation of reproductive cues to induce reproduction in ex situ systems14. Complementary to these active interventions are the recent recognition of the advantages of heterotrophic feeding in corals under high-temperature stress15 and the exploration of the role that food provision may play in reproduction16.
Heterotrophic feeding is known to influence the performance of corals17 and has been specifically linked to increased coral growth18,19, as well as thermal resistance and resilience20,21. Yet, the benefits of heterotrophy are not ubiquitous among coral species22 and can differ based on the type of food being consumed23, as well as the level of light exposure24. In the context of coral reproduction, heterotrophic feeding has shown variable results, with observations of higher25 as well as lower26 reproductive capacity following heterotrophic feeding being reported. The influence of heterotrophic feeding on coral reproduction across a spectrum of temperatures is rarely assessed, yet in the temperate coral Cladocora caespitosa, heterotrophy was found to be more important for reproduction under lower temperature conditions27. A better understanding of the role of temperature and feeding on reproductive output is likely needed to determine whether specific reefs (e.g., reefs associated with high food availability28) possess a higher capacity for recruitment under climate change.
Similar to reproductive output, the effect of temperature and feeding on reproductive timing in corals remains relatively understudied, despite the synchronization of reproduction with abiotic/biotic conditions being an important consideration for recruitment success in a warming ocean29. Warmer temperatures have been shown to result in earlier reproduction in coral thermal conditioning studies conducted in the lab30, and this has also been observed in corals collected from natural reefs across seasons31. Yet, interestingly the opposite trend was recently observed in fed corals cultured over the course of 1 year in an ex situ flow-through system (i.e., reproduction occurred earlier in the lunar cycle at cooler winter temperatures and later in the lunar cycle at warmer summer temperatures)32. This contrasting result suggests that reproductive timing may stray from typical patterns under conditions associated with abundant energetic resources.
Long-term controlled experiments under different temperature scenarios could contribute to a better understanding of the influence of heterotrophy on reproduction in scleractinian corals. Maintaining reproducing coral colonies under ex situ conditions for multiple reproductive cycles, however, can be challenging (but see previous research32,33). Herein, straightforward and effective techniques for the active feeding (food source: Artemia nauplii) and long-term culture of a brooding coral (Pocillopora acuta) in a flow-through aquaculture system are described; yet, it should be noted that all the techniques described can also be used in recirculating aquaculture systems. To demonstrate these techniques, a preliminary comparison of the reproductive output and timing of coral colonies held at 24 °C and 28 °C under "fed" and "unfed" treatments was conducted. These temperatures were chosen to approximate seawater temperatures in winter and summer, respectively, in southern Taiwan30,34; a higher temperature was not chosen because the promotion of long-term ex situ culture, rather than testing coral response to thermal stress, was a primary goal of this experiment. Further, the density of Artemia nauplii before and after the feeding sessions was quantified to compare the feasibility of heterotrophic feeding at both temperature treatments.
Specifically, 24 colonies of P. acuta (mean total linear extension ± standard deviation: 21.3 cm ± 2.8 cm) were obtained from flow-through tanks at the research facilities of the National Museum of Marine Biology & Aquarium, southern Taiwan. Pocillopora acuta is a common coral species that possesses both a broadcast spawning, but typically brooding reproductive strategy35,36. The parent colonies of these corals were originally collected from the Outlet reef (21.931°E, 120.745°N) approximately 2 years earlier for another experiment32. Consequently, the coral colonies used in the present experiment had been reared for their entire lives under ex situ culture conditions; specifically, the colonies were exposed to ambient temperature and a 12 h:12 h light: dark cycle at 250 µmol quanta m−2·s−1 and were fed Artemia nauplii twice per week. We recognize that this long-term ex situ culture could have affected how the colonies responded to the treatment conditions in this experiment. We, therefore, would like to emphasize that the primary aim here is to illustrate how the described techniques can be effectively used to culture corals ex situ by demonstrating an applied example wherein the effects of temperature and feeding on coral reproduction were assessed.
Coral colonies were evenly distributed across six flow-through system culture tanks (tank interior length x width x height: 175 cm x 62 cm x 72 cm; tank light regime: 12 h:12h light:dark cycle at 250 µmol quanta m−2·s−1) (Figure 1A). The temperature in three of the tanks was set at 28 °C, and the temperature in the other three tanks was set at 24 °C; each tank had a logger that recorded the temperature every 10 min (see the Table of Materials). The temperature was independently controlled in each tank using chillers and heaters, and water circulation was maintained using flow motors (see the Table of Materials). Half of the colonies in each tank (n = 2 colonies/tank) were fed Artemia nauplii twice per week, while the other colonies were not fed. Each feeding session was 4 h in duration and was conducted in two independent temperature-specific feeding tanks. During feeding, all the colonies were moved into the feeding tanks, including the unfed colonies, to standardize the potential stress effect of moving the colonies between the tanks. The colonies in the fed and unfed treatments were positioned in their own compartment using a meshed frame within the temperature-specific feeding tanks so that only the colonies in the fed condition received food. The coral reproductive output and timing were assessed for each colony daily at 09:00 A.M. by counting the number of larvae that had been released into the larval collection containers overnight.
Protocol
1. Hanging coral colonies in ex situ aquaculture tanks
- Position a notched bar (length x width x height: 75 cm x 1 cm x 3 cm), hereafter referred to as a "hanging bar", across the culture tank in preparation for hanging the coral colonies.
NOTE: The hanging bar used in this experiment was custom-made, but a simple PVC pipe with protruding screws (i.e., to act as the notches) would be sufficient as long as it can be positioned in a stable manner across the top of the culture tank and is strong enough to hold the corals. - Measure a piece of fishing line (see the Table of Materials) to ~1.5 m in length, and then fold it in half twice.
NOTE: The initial length of the fishing line should be chosen based on the desired final position of the coral colony in the culture tank. - Make a small overhand knot at the end of the folded fishing line that has the initial ends of the fishing line.
NOTE: After making the knot, there should be two large loops at the bottom and one small loop at the top. - Place the coral colony in the middle of the two large loops such that the loops are positioned around the colony and can securely hold the coral when it is hung in the water.
- Hook the small top loop of the fishing line into a notch on the hanging bar (Figure 1B).
2. Coral feeding
- Making the feeding container
- Construct a rectangular frame using acrylic pipe (length x width x height: 25 cm x 60 cm x 25 cm). Make two separate compartments in the frame where the fed and unfed corals can be placed, respectively (Figure 1C).
NOTE: Acrylic pipe was used because it is lightweight (i.e., as opposed to heavier PVC pipe) and, therefore, could facilitate easier movement of the feeding container in/out of the culture tanks. - Use a hot glue gun to adhere 100 µm of plankton mesh to the bottom and sides of the frame.
- Drill a total of ~10 small holes (0.5 cm in diameter) into the pipes (especially along the sides and the bottom of the frame) to prevent the feeding container from floating when placed in the culture tank.
- Drill holes (~0.5 cm in diameter) through the plankton mesh at each corner of the feeding container.
- Place an 8 cm length of 0.5 cm diameter tubing through the corner holes, and use a hot glue gun to fix it into position.
NOTE: These pieces of tubing will be connected to an air pump and bubble stones during feeding (see step 2.3.2 for more detail).
- Construct a rectangular frame using acrylic pipe (length x width x height: 25 cm x 60 cm x 25 cm). Make two separate compartments in the frame where the fed and unfed corals can be placed, respectively (Figure 1C).
- Artemia cultivation
- Collect 2 L of seawater from an independent feeding tank, and pour the seawater into an Artemia hatching container (Figure 1D).
NOTE: In the present experiment used to demonstrate the protocols, two independent treatment-specific feeding tanks were used, which necessitated the preparation of two hatching containers for Artemia cultivation. - Connect an air pump to tubing connected to the bottom of the hatching container for approximately 10 min prior to adding Artemia cysts.
- While waiting, use a balance to measure 8 g of Artemia cysts (see the Table of Materials).
NOTE: To obtain a mean density of 35 individual Artemia nauplii/mL, as suggested by Huang et al.19, use a ratio of 4 g of Artemia cysts to 1 L of seawater. - After 10 min, pour the 8 g of Artemia cysts into the hatching container.
- Incubate the Artemia cysts for 48 h.
- Collect 2 L of seawater from an independent feeding tank, and pour the seawater into an Artemia hatching container (Figure 1D).
- Preparing the feeding tank
- Place the feeding container into the feeding tank such that the top of the container is above the surface of the water.
- Connect the outer portion of the feeding container's corner tubing to an air pump, which will supply air to bubble stones for facilitating water circulation during feeding.
- Turn on the air pump ~5 min prior to the start of feeding.
- Artemia nauplii enrichment and collection
- Add 1.5 mL of enrichment diet (see the Table of Materials) to the hatching container 2 h before the desired feeding time.
NOTE: A ratio of 0.75 mL of enrichment diet to 1 L of seawater is recommended by Huang et al.19. - After 2 h, turn off the valve supplying air to the hatching container.
- Cover the hatching container with a cardboard box to exclude ambient light, and place a light source (a cell phone flashlight is sufficient) at the base of the hatching container for 5 min to attract Artemia nauplii to the bottom of the container and thereby facilitate the separation of live Artemia nauplii from empty shells.
- After 5 min, remove the box and the light source.
- Place a 3 L measuring jug below the hatching container.
- Detach the tubing from the hatching container to allow the Artemia nauplii and seawater solution to flow into the measuring jug; collect 1 L of the Artemia nauplii and seawater solution.
NOTE: Collect only half of the volume in the hatching container to exclude unwanted empty shells. - While standing in close proximity to the feeding tank, pour the Artemia nauplii and seawater solution through a 100 µm strainer to separate the Artemia nauplii (which will remain in the strainer) from the seawater.
- Rinse the Artemia nauplii held within the strainer twice with water from the feeding tank.
- The Artemia nauplii are now ready to be used.
- Add 1.5 mL of enrichment diet (see the Table of Materials) to the hatching container 2 h before the desired feeding time.
- Feeding the coral colonies
- Unload the Artemia nauplii by placing the strainer from step 2.4.8 into the feeding tank.
- Stir the water in the tank by hand to evenly distribute the Artemia nauplii.
NOTE: Collect samples for the "pre-feeding" quantification of the Artemia nauplii density after this step (see step 3.1 for more detail). - Move each hanging bar (with the coral colonies still hanging from the bar) from the culture tank to the feeding tank, and position the bar so that it is securely resting across the top of the feeding tank. The duration for which the corals are exposed to the air should be kept as short as possible.
NOTE: Make sure colonies are not touching each other and have enough space to capture food (e.g., ~5 cm apart). - Turn off the lights in the feeding tanks, or use a non-airtight lid to cover the feeding tank to avoid light disturbance during feeding.
- Allow the colonies to feed undisturbed for 4 h.
- After 4 h, collect the samples for the "post-feeding" quantification of the Artemia nauplii density (see step 3.1 for more detail).
- Post-feeding clean up
- After the feeding session is complete, remove the coral colonies. Take the hanging bars out of the feeding tank individually, and thoroughly rinse each coral with seawater from its respective culture tank to remove any residual Artemia nauplii.
NOTE: Rinse the colonies on a stable surface rather than while hanging to reduce the risk of damage that could occur if the colonies were to swing back and forth during rinsing. As per the initial transfer, keep the duration for which the corals are exposed to the air as short as possible. - Place the hanging bars (with corals hung) back into the culture tanks.
- Detach the tubes connecting the feeding container to the air pump, and remove the feeding container from the feeding tank.
- Rinse the feeding container thoroughly with fresh water to remove all the remaining Artemia nauplii.
- After the feeding session is complete, remove the coral colonies. Take the hanging bars out of the feeding tank individually, and thoroughly rinse each coral with seawater from its respective culture tank to remove any residual Artemia nauplii.
3. Quantifying Artemia nauplii density pre- and post-feeding
- Collecting the samples
- Collect samples at two time points: first, when the Artemia nauplii have been unloaded and distributed evenly in the feeding container (step 2.5.2), and again after the feeding session has been completed (step 2.5.6).
- For each time point, use three syringes to draw 20 mL of water from the surface, the middle layer, and the bottom layer of the feeding container, respectively.
- Sample dilution
- For each syringe, transfer the 20 mL of water sample into an independent 500 mL beaker.
- Add 180 mL of hot water (~60 °C) to the beaker (1:10 dilution).
NOTE: The hot water is used to immobilize the Artemia nauplii to increase the accuracy of enumeration. - Add 2 mL of the water sample from the beaker into each well of a 9-well plate.
NOTE: Mix the sample in the beaker to distribute the Artemia nauplii evenly in the water column before drawing the 2 mL of sample. - Count the number of Artemia nauplii in each well under a stereo microscope using 6.5x magnification (see the Table of Materials).
- Calculating the density of Artemia nauplii
- Divide the number of Artemia nauplii in each well by 2 to obtain the number of Artemia nauplii per mL. Then, multiply that number by 10 (to account for dilution) to calculate the Artemia nauplii density.
- Calculate the mean density of Artemia nauplii (i.e., average density across the 27 well replicates before vs. after feeding) to compare the Artemia nauplii density between pre- and post-feeding.
4. Coral larvae collection
- Making the larvae collection container (Figure 1E)
- Select a 6 L plastic water bottle, and cut the bottom of the bottle off completely.
NOTE: This opening will be used for transferring the colonies in and out of the larvae collection container. - Create two windows by cutting out a ~15 cm x 20 cm rectangle from each side of the bottle.
NOTE: A 6 L plastic water bottle is appropriate for corals that are ~15 cm in diameter; modify the size of the bottle based on the size of the corals being studied. - Use a hot glue gun and then epoxy to adhere a 100 µm plankton mesh onto each of the windows.
- Create two small holes (~0.5 cm in diameter) on each side of the bottom of the bottle.
- Put a string through the two small holes, and tie both ends to create a handle for hooking the larvae collection container onto the hanging bar.
- Before initial use, place the bottles into a flow-through tank (with no corals) for at least 24 h to remove any glue residue.
- Select a 6 L plastic water bottle, and cut the bottom of the bottle off completely.
- Preparing for coral collection
- Immerse the larvae collection container completely into the culture tank.
- Place the colony into the larvae collection container while keeping both the colony and the container submerged in water.
- Hook the handle of the larvae collection container onto the hanging bar.
NOTE: After hanging, ensure that the top of the collection container is ~3 cm above the water. - Repeat steps 4.2.1-4.2.3 until all the colonies are in their larvae collection containers.
- Collecting and enumerating the coral larvae
- Prepare a 3 L measuring jug, a bowl, a 3 mL pipette, and 50 mL tubes.
- Unhook the fishing line from the hanging bar, and remove one colony from its larvae collection container. Place the colony back into the culture tank immediately.
NOTE: Ensure that the duration of air exposure is as short as possible. - Place one hand on the cap end of the larvae collection container.
NOTE: When the larvae collection container is filled with water, it can be heavy. Without proper support, the container can break when it is being removed from the water. - Unhook the larvae collection container "handle" from the hanging bar.
- Slowly lift the larvae collection container out of the water.
- Hold the collection container at an approximately 45° angle above the culture tank for a few seconds to allow excess water to flow back into the tank via the larvae collection container windows.
NOTE: Do not angle the container past 45° to mitigate the chance of pouring out larvae from the top of the container. - Remove the larvae collection container from the tank, and position it on top of the measuring jug.
- Before unscrewing the cap, use one finger to apply a moderate amount of pressure against the cap, and then unscrew the cap.
NOTE: Water inside of the collection container can be released quickly when the cap is removed if it is not first supported by one's finger (i.e., potentially resulting in a loss of larvae). - Transfer some of the water inside of the measuring jug into a bowl.
- Manually count the number of larvae in the bowl by using a 3 mL pipette to move the larvae into a 50 mL tube.
NOTE: Be aware that some of the larvae may get stuck inside the pipette. If this happens, draw some seawater into the pipette, and shake gently while sealing the pipette with one finger to loosen the larvae. - Continue step 4.3.9 and step 4.3.10 until all the larvae have been counted. At this stage, the larvae can be used in subsequent experiments.
- Repeat steps 4.3.2-4.3.10 for all the other coral colonies.
NOTE: The measuring jug and bowl should be rinsed between colonies. - After the counting is finished, rinse each collection container thoroughly with fresh water, especially the windows.
Representative Results
The described protocols allowed for (1) the comparison of the reproductive output and timing of individual coral colonies among distinct feeding and temperature treatments and (2) an assessment of the feasibility of Artemia nauplii feeding at different temperatures. Herein, a brief overview of the findings is given, but caution should be exercised with regard to the broad interpretation of the reported effects of temperature and feeding on coral reproduction due to the short-term nature of this experiment (i.e., only one reproductive cycle) and the use of coral colonies acclimated to ex situ conditions.
Each colony reproduced over the course of our monitoring period (lunar September 2022), and the total monthly reproductive output showed high variation among the colonies. The total number of larvae released by colonies ranged from 6 to 319, except for one colony (in the unfed 24 °C treatment) that produced 528 larvae; data for all the colonies are shown in Figure 2, but the high-producing outlier colony was not included in the data analysis. Reproductive output was affected by temperature (generalized linear mixed-effects model; z = 5.35, p < 0.001) and feeding (z = 3.01, p < 0.003), with a significant interaction found between the temperature and feeding treatments (z = 12.22, p < 0.001). Colonies cultured at 28 °C released more larvae when unfed (mean ± standard deviation; 151 ± 82) than when fed (131 ± 133) (generalized linear mixed-effects model, post hoc contrast; z = 3.01, p = 0.014), but the opposite trend was found in colonies cultured at 24 °C, whereby the fed colonies (80 ± 78) produced more larvae than the unfed colonies (12 ± 6) (z = 11.91, p < 0.001).
Reproduction in all the colonies occurred before the full moon (lunar day 15) (Figure 3). The mean lunar day (MLD) of larval release (weighted by reproductive output) ranged from lunar day 6.5 to lunar day 11.1, with a significant difference among treatments only being detected between the "unfed 28 °C" colonies, which reproduced earlier in the lunar cycle, and the "fed 24 °C" colonies, which reproduced later in the lunar cycle (linear mixed-effects model, post hoc contrast, t = 4.10, p = 0.006).
In the month prior to formal reproduction monitoring (lunar August 2022), the Artemia nauplii density was assessed before and after feeding sessions; this was repeated at three time points: at the onset of coral culture for this experiment (T0) and 2 weeks and 4 weeks into coral culture under treatment conditions (Figure 4). The initial assessment at T0 showed no difference between the pre- and post-feeding density of Artemia nauplii in both temperature treatments. After 2 weeks and 4 weeks of culture, the Artemia nauplii density was lower after feeding in both temperature treatments (week 2: two-way ANOVA, F1,104 = 128.45, p < 0.001; week 4: two-way ANOVA, F1,104 = 294.71, p < 0.001). There was no difference in the pre-feeding density between temperature treatments (p > 0.05) or the post-feeding density between temperature treatments (p > 0.05) at any of the three time points assessed.
All the analyses were conducted in R using the packages lme437, lmerTest38, emmeans39, car40, and Hmisc41. The data and R script used for the analyses are publicly available on GitHub (https://github.com/CJ-McRae/Lam-et-al_JoVE-submission).
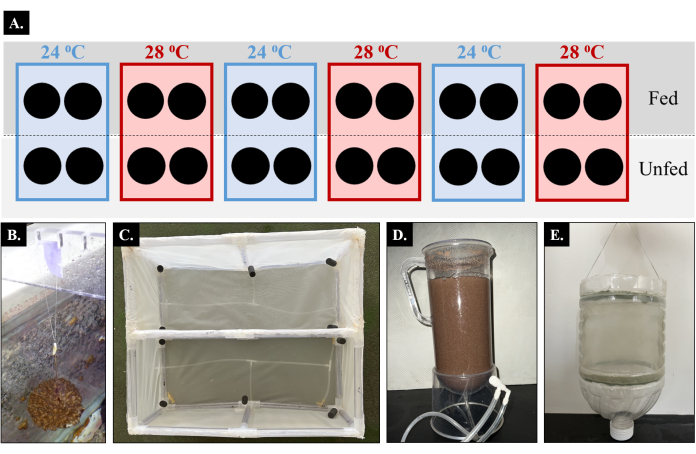
Figure 1: Schematic of experimental design and representative materials for the feeding and ex situ culture of a brooding scleractinian coral. (A) Colonies of Pocillopora acuta were cultured in flow-through aquaculture tanks at either 24 °C or 28 °C under fed and unfed conditions; the black circles represent the colonies. (B) The colonies were hung with fishing lines to reduce handling stress and to promote efficient movement between the culture and feeding tanks. (C) During feeding sessions, all the colonies were moved into a meshed frame within temperature-specific feeding tanks. Fed colonies were positioned in one compartment of the frame, and unfed colonies were positioned in the other compartment of the frame; only fed colonies were provided with food. (D) Enriched Artemia nauplii were given to the colonies in the fed treatment twice per week. (E) The colonies were put into larvae collection containers overnight to quantify the reproductive output daily over the course of one lunar cycle. Please click here to view a larger version of this figure.
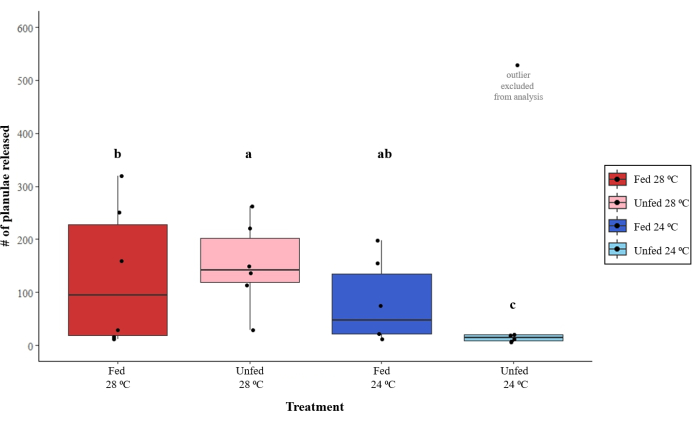
Figure 2: Reproductive output of Pocillopora acuta colonies under different temperatures (24 °C vs. 28 °C) and feeding treatments (fed vs. unfed). The letters are representative of significant differences in reproductive output among treatments. Please click here to view a larger version of this figure.
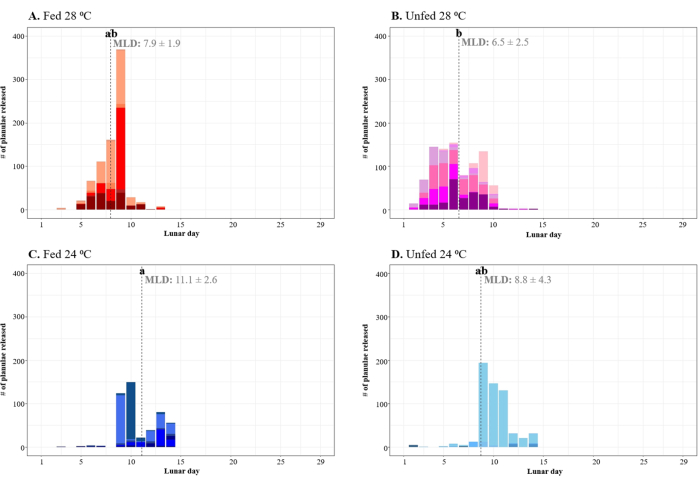
Figure 3: Reproductive timing of Pocillopora acuta colonies under different temperatures (24 °C vs. 28 °C) and feeding treatments (fed vs. unfed). The vertical dashed line shows the mean lunar day (MLD) of reproduction for each treatment. The color tones within each bar of the treatment-specific plots (A–D) indicate the contribution of individual colonies to the daily total reproduction. The letters are representative of significant differences in reproductive timing among treatments. Please click here to view a larger version of this figure.
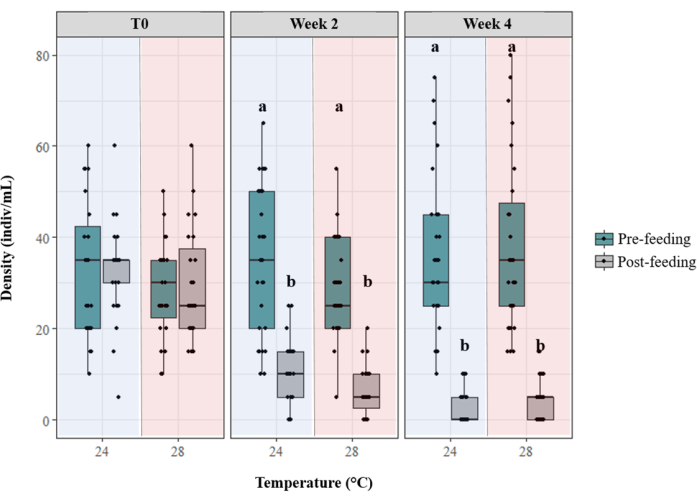
Figure 4: Density of Artemia nauplii before and after coral feeding sessions within the 24 °C and 28 °C temperature treatments. The pre-feeding density was calculated prior to coral feeding, and the post-feeding density was calculated after the completion of a 4 h coral feeding session. The density of Artemia nauplii was assessed at the beginning of coral culture (T0) and then after 2 weeks and 4 weeks under treatment conditions in a flow-through aquaculture system. Please click here to view a larger version of this figure.
Discussion
This preliminary assessment of the effect of temperature and feeding on coral reproduction revealed differences in reproductive output and timing among colonies cultured under distinct treatment conditions. Further, it was found that feeding Artemia nauplii to coral colonies appeared to be effective at relatively cool (24°C) as well as warm temperatures (28 °C). These combined findings highlight the applicability of these straightforward techniques for the feeding and culture of reproducing scleractinian corals (using P. acuta as an example) in ex situ aquaculture systems.
In the context of reproductive output, feeding was found to have a differing influence depending on the temperature treatment in which the colonies were cultured, whereby feeding only appeared to have a positive effect on reproductive output in colonies held in the 24 °C treatment. This result is somewhat surprising since in other marine organisms, limited food provision at high temperatures has shown a negative effect on reproduction (e.g., reduction in spawning in damselfish42) and has been associated with poor early life stage development (e.g., higher mortality and reduced growth in crabs during metamorphosis43). In corals, specific assessments of the interactive effects of feeding and temperature have been primarily focused on the photochemical performance of the coral's algal symbionts44,45, and these interactive effects are rarely explored in the context of reproduction. Future studies targeting a comprehensive assessment of the reproduction-based effects of feeding at different temperatures over multiple reproductive cycles are needed. This, however, was not the goal of the present experiment. Instead, this experiment was primarily used to demonstrate the efficacy of the presented feeding and culture techniques. Through the use of these techniques, clear reproductive trends of individual colonies can be easily assessed, which is important, as inter-colony variation in reproductive output is not uncommon. For example, a wide range of reproductive output has been found among colonies, as well as over time for the same individual colony, in multiple studies30,32,46,47. Possible explanations for the high variability in reproductive output include plasticity in reproductive strategies and/or shifts in the prioritization of energy allocation48,49. Techniques that allow for colony-specific assessments of reproductive output, such as those described in this experiment, can help identify the environmental/genetic drivers of reproductive capacity, pertinent to our understanding of coral recruitment (i.e., relevant to natural reef resilience) and broodstock supply potential (i.e., relevant to ex situ cultivation aimed at supporting coral restoration).
The assessment of reproductive timing in this experiment revealed that only the colonies in the "unfed 28 °C" treatment released larvae significantly earlier than colonies in the "fed 24 °C" treatment; the timing remained similar among the other treatments. Temperature-driven plasticity in reproductive timing has been observed across multiple coral species, with advanced timing observed at warmer temperatures50,51,52. This shift in timing is likely explained by the accelerated development of the gametes and embryos at warmer temperatures53, which under climate change, could ultimately have either an adaptive or disruptive influence on coral reproduction and recruitment54,55,56. Experiments that explicitly examine the possible interactive relationship between feeding and temperature on reproductive timing could provide a better understanding of the consequences of timing shifts and could also test the practicality of increasing the frequency of reproductive cycles to enhance ex situ aquaculture production.
Effective ex situ feeding techniques are required to conduct controlled experiments exploring the potential interactive relationship between temperature and feeding on coral reproduction. In this experiment, coral colonies were fed in temperature-specific feeding tanks at 24 °C and 28 °C, and similar patterns in the pre- and post-feeding density of Artemia nauplii were found across the temperature treatments (i.e., lower density of Artemia nauplii post-feeding vs. pre-feeding). This is indicative of three important points: (1) the temperature treatment did not appear to affect the health of the Artemia nauplii; (2) the coral colony feeding rates were approximately the same at both temperatures; and (3) the coral colonies consumed Artemia nauplii during feeding sessions at both temperatures (except for the T0 time point, which may be indicative of colony stress when acclimating to the experimental conditions). It is important to point out that the interpretation of the density trends between the temperature treatments and across time serves only as a proxy-based assessment. Robust investigation to confirm feeding (e.g., examination of gut content57) and Artemia nauplii physiology (e.g., heat shock protein expression58) would be needed to make definitive conclusions about the feeding feasibility; an assessment of this nature was outside of the scope of this experiment. Yet, based on the experiment data coupled with visual confirmation during feeding, we are confident that the coral colonies in this experiment were actively feeding under both treatment temperatures. Corals can show contrasting responses to feeding at high temperatures, with some species showing a reduction and others showing an increase in feeding rate45. Therefore, species- and location-specific temperature tolerance should be taken into consideration when determining feeding temperatures in future experiments.
The feeding and culture techniques described provide several advantages that seek to improve both the quality of coral health and the longevity of reproduction in ex situ aquaculture. The overarching goal guiding this described approach was founded on minimizing potential sources of coral stress. To begin, the need for direct handling of the coral colonies was eliminated by using fishing lines to hang the corals. This facilitates the efficient movement of colonies between culture and feeding tanks and allows for simple and fast adjustments to the colony position (e.g., shortening or lengthening the fishing line to change the depth of the colony in the tank). In contrast to colonies being placed on a stand or at the bottom of a culture tank, hanging the colonies promotes growth in all dimensions, reduces the accumulation of algae, and creates more useable space in the tank (e.g., multiple corals can be suspended vertically on a single fishing line if needed)59. In addition to reducing the need for energetic trade-offs60, which can help promote long-term reproduction32, the feeding techniques described also helped reduce coral stress. Feeding in independent tanks is recommended19,61 (as opposed to directly in the culture tanks) to mitigate exposure of the corals to potentially high nutrient levels, which can be detrimental to coral health and lead to abundant algal growth62,63,64,65. Further, if water quality issues do arise, maintaining and changing the water within the feeding tanks can be done easily without disturbing the coral colonies. The larvae collection containers were also designed with coral stress reduction in mind, whereby colony-specific reproduction could be achieved without direct handling or the need for culture in single-colony tanks. Having multiple colonies in large culture tanks may help improve reproductive longevity (especially in coral species with mixed reproductive modes49), which has been shown to decline over time in ex situ systems66,67. In addition, the use of large plastic bottles as larvae collection containers provides the larvae with abundant space, which can reduce settlement on the collection container itself; fast settlement can be problematic when small collection containers are used (McRae and Lam, personal observations). Lastly, these ex situ feeding and culture techniques use materials that are cost-effective, easy to make, and can be customized according to experiment-specific needs.
The main limitations of the described feeding and culture techniques include 1) a finite limit on the number of colonies that can be cultured due to tank space requirements, 2) the inability to standardize the reproductive mode (sexual vs. asexual) due to multiple colonies being cultured within the same tank, and 3) the use of a single species from a single reef site to test the efficacy of the described techniques. Future research would benefit from testing how other coral species perform using these feeding and culture techniques, as well as exploring the use of other food types to best cater to species-specific dietary needs.
In conclusion, it is recognized that critiques of other active interventions68,69,70 are likely also applicable to the promotion of ex situ culture for reproducing corals, as the key limitations (e.g., scalability, genetic diversity) remain relevant. Yet, similar to other active interventions, ex situ coral culture is not intended to be viewed as a singular solution but rather as a supportive approach that should be explored in tandem with meaningful climate change mitigations. Through the use of the described techniques, coral stress can be reduced to improve the reproductive longevity of a scleractinian coral, P. acuta, in ex situ aquaculture systems, from which the colonies (and their offspring) can contribute to research and restoration efforts.
Declarações
The authors have nothing to disclose.
Acknowledgements
This research was funded by the Ministry of Science and Technology (Taiwan), grant numbers MOST 111-2611-M-291-005 and MOST 111-2811-M-291-001.
Materials
| Artemia cysts | Supreme plus | NA | Food source |
| Chiller | Resun | CL650 | To cool down water temperature if needed |
| Conductivity portable meter | WTW | Cond 3110 | To measure salinity |
| Enrichment diets | Omega | NA | Used in Artemia cultivation |
| Fishing line | Super | Nylon monofilament | To hang the coral colonies |
| Flow motors | Maxspect | GP03 | To create water flow |
| Heater 350 W | ISTA | NA | Heaters used in tanks |
| HOBO pendant temperature logger | Onset Computer | UA-002-08 | To record water temperature |
| LED lights | Mean Well | FTS: HLG-185H-36B | NA |
| Light portable meter | LI-COR | LI-250A | Device used with light sensor to measure light intensity in PAR |
| Light sensor | LI-COR | LI-193SA | NA |
| Plankton net 100 µm mesh size | Omega | NA | To collect larvae and artemia |
| Primary pump 6000 L/H | Mr. Aqua | BP6000 | To draw water from tanks into chiller |
| Propeller-type current meter | KENEK | GR20 | Device used with propeller-type detector to measure flow rate |
| Propeller-type detector | KENEK | GR3T-2-20N | NA |
| Stereo microscope | Zeiss | Stemi 2000-C | To count the number of artemia |
| Temperature controller 1000 W | Rep Park | O-RP-SDP-1 | To set and maintain water temperature |
Referências
- Hughes, T. P., et al. Coral reefs in the Anthropocene. Nature. 546 (7656), 82-90 (2017).
- Special Report on the Ocean and Cryosphere in a changing climate. Intergovernmental Panel on Climate Change Available from: https://www.ipcc.ch/srocc/ (2019)
- van Oppen, M. J. H., Lough, J. M. Synthesis: Coral bleaching: patterns, processes, causes and consequences. Coral Bleaching: Patterns, Processes, Causes and Consequences. , 343-348 (2018).
- Glynn, P. W. Coral reef bleaching: Ecological perspectives. Coral Reefs. 12 (1), 1-17 (1993).
- Hughes, T. P., et al. Spatial and temporal patterns of mass bleaching of corals in the Anthropocene. Science. 359 (6371), 80-83 (2018).
- Grottoli, A. G., et al. The cumulative impact of annual coral bleaching can turn some coral species winners into losers. Global Change Biology. 20 (12), 3823-3833 (2014).
- Frieler, K., et al. Limiting global warming to 2 °C is unlikely to save most coral reefs. Nature Climate Change. 3 (2), 165-170 (2013).
- Montefalcone, M., Morri, C., Bianchi, C. N. Long-term change in bioconstruction potential of Maldivian coral reefs following extreme climate anomalies. Global Change Biology. 24 (12), 5629-5641 (2018).
- Traylor-Knowles, N. Heat stress compromises epithelial integrity in the coral, Acropora hyacinthus. PeerJ. 7, e6510 (2019).
- Anthony, K. R. N., Hoogenboom, M. O., Maynard, J. A., Grottoli, A. G., Middlebrook, R. Energetics approach to predicting mortality risk from environmental stress: a case study of coral bleaching. Functional Ecology. 23 (3), 539-550 (2009).
- Ward, S., Harrison, P., Hoegh-Guldberg, O. Coral bleaching reduces reproduction of scleractinian corals and increases susceptibility to future stress. Proceedings of the 9th Coral Reef Symposium. , 1123-1128 (2002).
- Suzuki, G., et al. Enhancing coral larval supply and seedling production using a special bundle collection system "coral larval cradle" for large-scale coral restoration. Restoration Ecology. 28 (5), 1172-1182 (2020).
- Schmidt-Roach, S., et al. Novel infrastructure for coral gardening and reefscaping. Frontiers in Marine Science. 10, 1110830 (2023).
- Craggs, J., et al. Inducing broadcast coral spawning ex situ: Closed system mesocosm design and husbandry protocol. Ecology and Evolution. 7 (24), 11066-11078 (2017).
- Conti-Jerpe, I. E., et al. Trophic strategy and bleaching resistance in reef-building corals. Science Advances. 6 (15), 5443 (2020).
- Bellworthy, J., Spangenberg, J. E., Fine, M. Feeding increases the number of offspring but decreases parental investment of Red Sea coral Stylophora pistillata. Ecology and Evolution. 9 (21), 12245-12258 (2019).
- Houlbrèque, F., Ferrier-Pagès, C. Heterotrophy in tropical scleractinian corals. Biological Reviews. 84 (1), 1-17 (2009).
- Ferrier-Pagès, C., Witting, J., Tambutté, E., Sebens, K. P. Effect of natural zooplankton feeding on the tissue and skeletal growth of the scleractinian coral Stylophora pistillata. Coral Reefs. 22 (3), 229-240 (2003).
- Huang, Y. -. L., Mayfield, A. B., Fan, T. -. Y. Effects of feeding on the physiological performance of the stony coral Pocillopora acuta. Scientific Reports. 10 (1), 19988 (2020).
- Tagliafico, A., et al. Lipid-enriched diets reduce the impacts of thermal stress in corals. Marine Ecology Progress Series. 573, 129-141 (2017).
- Huffmyer, A. S., Johnson, C. J., Epps, A. M., Lemus, J. D., Gates, R. D. Feeding and thermal conditioning enhance coral temperature tolerance in juvenile Pocillopora acuta. Royal Society Open Science. 8 (5), 210644 (2021).
- Grottoli, A. G., Rodrigues, L. J., Palardy, J. E. Heterotrophic plasticity and resilience in bleached corals. Nature. 440 (7088), 1186-1189 (2006).
- Conlan, J. A., Bay, L. K., Severati, A., Humphrey, C., Francis, D. S. Comparing the capacity of five different dietary treatments to optimise growth and nutritional composition in two scleractinian corals. PLoS One. 13 (11), 0207956 (2018).
- Treignier, C., Grover, R., Ferrier-Pagés, C., Tolosa, I. Effect of light and feeding on the fatty acid and sterol composition of zooxanthellae and host tissue isolated from the scleractinian coral Turbinaria reniformis. Limnology and Oceanography. 53 (6), 2702-2710 (2008).
- Gori, A., et al. Effects of food availability on the sexual reproduction and biochemical composition of the Mediterranean gorgonian Paramuricea clavata. Journal of Experimental Marine Biology and Ecology. 444, 38-45 (2013).
- Séré, M. G., Massé, L. M., Perissinotto, R., Schleyer, M. H. Influence of heterotrophic feeding on the sexual reproduction of Pocillopora verrucosa in aquaria. Journal of Experimental Marine Biology and Ecology. 395 (1), 63-71 (2010).
- Rodolfo-Metalpa, R., Peirano, A., Houlbrèque, F., Abbate, M., Ferrier-Pagès, C. Effects of temperature, light and heterotrophy on the growth rate and budding of the temperate coral Cladocora caespitosa. Coral Reefs. 27 (1), 17-25 (2008).
- Fox, M. D., et al. Gradients in primary production predict trophic strategies of mixotrophic corals across spatial scales. Current Biology. 28 (21), 3355-3363 (2018).
- Shlesinger, T., Loya, Y. Breakdown in spawning synchrony: A silent threat to coral persistence. Science. 365 (6457), 1002-1007 (2019).
- McRae, C. J., Huang, W. -. B., Fan, T. -. Y., Côté, I. M. Effects of thermal conditioning on the performance of Pocillopora acuta adult coral colonies and their offspring. Coral Reefs. 40 (5), 1491-1503 (2021).
- Fan, T. Y., et al. Plasticity in lunar timing of larval release of two brooding pocilloporid corals in an internal tide-induced upwelling reef. Marine Ecology Progress Series. 569, 117-127 (2017).
- Lam, K. -. W., et al. Consistent monthly reproduction and completion of a brooding coral life cycle through ex situ culture. Diversity. 15 (2), 218 (2023).
- O’Neil, K. L., Serafin, R. M., Patterson, J. T., Craggs, J. R. K. Repeated ex situ Spawning in two highly disease susceptible corals in the family Meandrinidae. Frontiers in Marine Science. 8, 669976 (2021).
- Keshavmurthy, S., et al. Coral Reef resilience in Taiwan: Lessons from long-term ecological research on the Coral Reefs of Kenting national park (Taiwan). Journal of Marine Science and Engineering. 7 (11), 388 (2019).
- Smith, H. A., Moya, A., Cantin, N. E., van Oppen, M. J. H., Torda, G. Observations of simultaneous sperm release and larval planulation suggest reproductive assurance in the coral Pocillopora acuta. Frontiers in Marine Science. 6, 362 (2019).
- Yeoh, S. -. R., Dai, C. -. F. The production of sexual and asexual larvae within single broods of the scleractinian coral, Pocillopora damicornis. Marine Biology. 157 (2), 351-359 (2010).
- Bates, D., Mächler, M., Bolker, B., Walker, S. Fitting linear mixed-effects models using lme4. Journal of Statistical Software. 67 (1), 1-48 (2015).
- Kuznetsova, A., Brockhoff, P. B., Christensen, R. H. B. lmerTest package: Tests in linear mixed effects models. Journal of Statistical Software. 82 (13), 1-26 (2017).
- Length, R. . Emmeans: Estimated marginal means, aka least-squares means. R Package Version 1.7.4-1. , (2022).
- Fox, J., Weisberg, S. . An R Companion to Applied Regression. Third edition. , (2019).
- Harell, F. E. . Hmisc: Harrell Miscellaneous_. R package version 4.7-1. , (2022).
- Donelson, J. M., Munday, P. L., McCormick, M. I., Pankhurst, N. W., Pankhurst, P. M. Effects of elevated water temperature and food availability on the reproductive performance of a coral reef fish. Marine Ecology Progress Series. 401, 233-243 (2010).
- Torres, G., Giménez, L. Temperature modulates compensatory responses to food limitation at metamorphosis in a marine invertebrate. Functional Ecology. 34 (8), 1564-1576 (2020).
- Borell, E. M., Bischof, K. Feeding sustains photosynthetic quantum yield of a scleractinian coral during thermal stress. Oecologia. 157 (4), 593-601 (2008).
- Ferrier-Pagès, C., Rottier, C., Beraud, E., Levy, O. Experimental assessment of the feeding effort of three scleractinian coral species during a thermal stress: Effect on the rates of photosynthesis. Journal of Experimental Marine Biology and Ecology. 390 (2), 118-124 (2010).
- Harriott, V. J. Reproductive seasonality, settlement, and post-settlement mortality of Pocillopora damicornis (Linnaeus), at Lizard Island, Great Barrier Reef. Coral Reefs. 2 (3), 151-157 (1983).
- Shefy, D., Shashar, N., Rinkevich, B. The reproduction of the Red Sea coral Stylophora pistillata from Eilat: 4-decade perspective. Marine Biology. 165 (2), 27 (2018).
- Rinkevich, B., Loya, Y. Variability in the pattern of sexual reproduction of the coral Stylophora pistillata at Eilat, Red Sea: a long-term study. The Biological Bulletin. 173 (2), 335-344 (1987).
- Combosch, D. J., Vollmer, S. V. Mixed asexual and sexual reproduction in the Indo-Pacific reef coral Pocillopora damicornis. Ecology and Evolution. 3 (10), 3379-3387 (2013).
- Fan, T. -. Y., Dai, C. -. F. Reproductive plasticity in the reef coral Echinopora lamellosa. Marine Ecology Progress Series. 190, 297-301 (1999).
- Crowder, C. M., Liang, W. -. L., Weis, V. M., Fan, T. -. Y. Elevated temperature alters the lunar timing of planulation in the brooding Coral Pocillopora damicornis. PLoS One. 9 (10), e107906 (2014).
- Lin, C. -. H., Nozawa, Y. The influence of seawater temperature on the timing of coral spawning. Coral Reefs. 42, 417-426 (2023).
- O’Connor, M. I., et al. Temperature control of larval dispersal and the implications for marine ecology, evolution, and conservation. Proceedings of the National Academy of Sciences of the United States of America. 104 (4), 1266-1271 (2007).
- Nozawa, Y. Annual variation in the timing of coral spawning in a high-latitude environment: Influence of temperature. The Biological Bulletin. 222 (3), 192-202 (2012).
- Bouwmeester, J., et al. Solar radiation, temperature and the reproductive biology of the coral Lobactis scutaria in a changing climate. Scientific Reports. 13 (1), 246 (2023).
- Bouwmeester, J., et al. Latitudinal variation in monthly-scale reproductive synchrony among Acropora coral assemblages in the Indo-Pacific. Coral Reefs. 40 (5), 1411-1418 (2021).
- Lai, S., et al. First experimental evidence of corals feeding on seagrass matter. Coral Reefs. 32 (4), 1061-1064 (2013).
- Iryani, M. T. M., et al. Cyst viability and stress tolerance upon heat shock protein 70 knockdown in the brine shrimp Artemia franciscana. Cell Stress and Chaperones. 25 (6), 1099-1103 (2020).
- Nedimyer, K., Gaines, K., Roach, S. Coral Tree Nursery©: An innovative approach to growing corals in an ocean-based field nursery. Aquaculture, Aquarium, Conservation & Legislation. 4, 442-446 (2011).
- Leuzinger, S., Willis, B. L., Anthony, K. R. N. Energy allocation in a reef coral under varying resource availability. Marine Biology. 159 (1), 177-186 (2012).
- Chang, T. C., Mayfield, A. B., Fan, T. Y. Culture systems influence the physiological performance of the soft coral Sarcophyton glaucum. Science Reports. 10 (1), 20200 (2020).
- Forsman, Z. H., Kimokeo, B. K., Bird, C. E., Hunter, C. L., Toonen, R. J. Coral farming: Effects of light, water motion and artificial foods. Journal of the Marine Biological Association of the United Kingdom. 92 (4), 721-729 (2012).
- Costa, A. P. L., et al. The effect of mixotrophy in the ex situ culture of the soft coral Sarcophyton cf. glaucum. Aquaculture. 452, 151-159 (2016).
- Marubini, F., Davies, P. S. Nitrate increases zooxanthellae population density and reduces skeletogenesis in corals. Marine Biology. 127 (2), 319-328 (1996).
- Bartlett, T. C. Small scale experimental systems for coral research: Considerations, planning, and recommendations. NOAA Technical Memorandum NOS NCCOS 165 and CRCP 18. , 68 (2013).
- Galanto, N., Sartor, C., Moscato, V., Lizama, M., Lemer, S. Effects of elevated temperature on reproduction and larval settlement in Leptastrea purpurea. Coral Reefs. 41 (2), 293-302 (2022).
- Nietzer, S., Moeller, M., Kitamura, M., Schupp, P. J. Coral larvae every day: Leptastrea purpurea, a brooding species that could accelerate coral research. Frontiers in Marine Science. 5, 466 (2018).
- Edwards, A. J., et al. Direct seeding of mass-cultured coral larvae is not an effective option for reef rehabilitation. Marine Ecology Progress Series. 525, 105-116 (2015).
- Boström-Einarsson, L., et al. Coral restoration – A systematic review of current methods, successes, failures and future directions. PLoS One. 15 (1), 0226631 (2020).
- Anthony, K. R. N., et al. Interventions to help coral reefs under global change-A complex decision challenge. PLoS One. 15 (8), e0236399 (2020).
