All protocols described in this manuscript for blood collection, isolation and analysis have been reviewed and approved by the Institutional Review Board at the University of Giessen, Germany. The consent of the patients to include their samples in the study was obtained. All steps for isolation and cell culture are carried out under a biological safety cabinet.
1. Venipuncture
- Prepare all equipment needed for blood collection including disinfection spray, sterile swab, blood collection cannula with 80 mm tube and multi-adapter, tourniquet/blood pressure cuff, Monovette 9 mL lithium-heparin.
NOTE: EDTA as an anticoagulant is also effective. - Collect blood from the most appropriate arm vein, usually vena mediana cubiti or vena cephalica.
- Apply a tourniquet/blood pressure cuff with light pressure, around 80 mm/Hg.
- Disinfect gloves and puncture site with disinfectant spray containing alcohol. Allow the disinfected puncture site to air dry.
- The veins protrude due to the pressure of the pressure cuff. Insert the needle (Cannula diameter (outer) 21G / 0.8 mm, length 19 mm) at a 15°-20° angle of the vein attempting to avoid trauma and minimizing probing.
- Take blood with appropriate system, 4 tubes containing 9 mL of blood (more than 7-8 tubes are problematic for one experimenter to isolate properly).
- After blood collection, place the collection tubes in the dark for 5 min to ensure uniform anticoagulation.
2. PBMC isolation
- Prepare all needed solutions as described below.
- Bring Dulbecco's balanced salt solution (DPBS; concentration 1x) and lymphocyte isolation medium (1.077 g/mL) to room temperature (20-25 °C).
- Prepare Fetal bovine serum (FBS) at room temperature and keep one sterile 50 mL conical tube with FBS on ice. For each blood sample, 2 mL of FBS is required.
- Store freezing container at 4 °C and precool cryotubes at 4 °C.
- Warm cell culture medium to 37 °C, the medium consists of RPMI 1640 with 50 mL of FBS and penicillin 50 U/mL streptomycin 50 U/mL. This solution can be stored at 3 °C for up to 2 months.
- Add 8 mL of DPBS in sterile 50 mL conical tubes. Add 15 mL of lymphocyte isolation medium in sterile 50 mL conical tube (Medium is light sensitive, add it before starting isolation).
- Add 8 mL of blood to 8 mL of DPBS, and carefully further mix with a 3 mL plastic Pasteur pipette.
- Layer the blood/DPBS mix gently with a 3 mL plastic Pasteur pipette on top of the lymphocyte isolation medium. To apply the first layer to the medium, tilt the tube 20°-30°, which will result in less penetration of the blood-PBS mixture into the medium layer.
- Layer the blood-PBS mixture carefully over the side wall of the tube onto the lymphocyte isolation medium. Use a steady speed to keep the blood flow constant.
- In the next step, bring the tube slowly into an upright position, the remaining blood is carefully layered over the side wall of the tube onto the blood layer.
- Centrifuge for 10 min at 1000 x g at room temperature in a centrifuge with a swinging-bucket rotor with brakes off. After centrifugation, the blood/PBMC mix is separated into four layers. The top layer consists of plasma and platelets, the second layer is the PBMC layer, followed by a lymphocyte isolation medium layer, and, finally, erythrocytes and granulocytes in the bottom layer. The different layers are displayed in Figure 1.
- Remove 2/3rd of the plasma layer with a plastic Pasteur pipette.
- Using a 1 mL pipette, collect the PBMCs on the lymphocyte isolation medium layer taking care not to get any medium in the sample.
- Place the tip 1 mm above the PBMC layer. The PBMC layer should not be punctured, otherwise the medium will flow over the cells. The suction of the pipette pulls the PBMCs to this point so that they can be collected several times at this point.
NOTE: To maximize the number of PBMCs collected, at the end of the procedure search for the rest on the surface and try to collect the cells there as well. To stabilize the procedure, the tube can be placed on a surface. - Transfer the PBMCs step by step to a new 50 mL tube until the layer is completely harvested. Add DPBS up to the 25 mL mark, wash away lymphocyte isolation medium and other residues.
- Centrifuge for 10 min at 100 x g at room temperature with brakes on. Remove the supernatant with a vacuum pump or similar, take care not to damage the cell pellet.
- Resuspend the pellet in 1 mL of DPBS and add up DPBS to the 25 mL mark. Repeat washing once again and then resuspend in medium appropriate for the next steps.
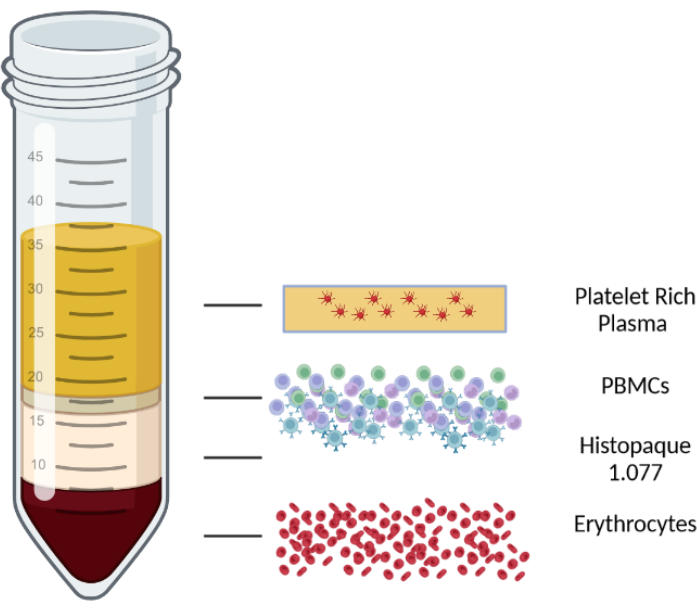
Figure 1: Schematic representation of a density gradient centrifugation to illustrate the different layers. Please click here to view a larger version of this figure.
3. Cryopreservation
- Cool down a freezing container to 4 °C, cool FBS on ice.
- Resuspend PBMC pellet in 1 mL of FBS with a 1 mL pipette, the FBS should be at room temperature.
- Mix DMSO with pre-cooled FBS on ice 1:5, final concentration 20% DMSO then put the FBS: DMSO mixture back on ice. Always prepare the solution fresh.
- Transfer the PBMC cell suspension in FBS to well labelled 2 mL cryotubes. Use one cryotube per collection tube. Adjust the number of cells between 1 x 107 and 5 x 107 per mL. Use an automated cell counter to determine cell count and viability.
- Add 1 mL of FBS: DMSO mixture dropwise with a 1 mL pipette, approximately 1-2 drops per s, into the tube. The dropwise addition leads to continuous and consistent mixing.
- Place tubes in the precooled freezing container. Place the freezing container in a -80 °C freezer for 24 h. The freezing container provides a controlled cooling of -1 °C per min.
- Remove the tubes from the -80 °C freezer. After removal from the freezer store the tubes in the gas phase of liquid nitrogen. Document the place of each sample.
4. Thawing
- Prepare all needed solutions: Cell culture medium RPMI 1640 with 50 mL of FBS and penicillin 50 U/mL, streptomycin 50 U/mL. This solution can be stored at 3°C for up to 2 months.
- Pre-warm cell culture medium to 37 °C. Add 3 mL of pre-warm cell culture medium into sterile 50 mL conical tubes.
- Remove sample from liquid nitrogen tank. Thaw samples in a water bath at 37 °C for approx. 3.5 min, remove from the water bath as soon as the last ice is melting. A pinhead-sized piece of ice should still be visible in the tube.
NOTE: DMSO is harmful to PBMCs work as quickly as possible. - Remove the cell:FBS:DMSO mix with a 1 mL pipette from the cryotube. Mix the PBMC samples with the cell culture medium in the prepared 50 mL tubes. Wash out the tubes with 5 mL of culture medium in three steps of 2 mL, 2 mL, and 1 mL.
- Transfer the medium into the tubes. This is performed to transfer possible cell residues. Centrifuge for 10 min at 100 x g at room temperature.
- Discard the supernatant and add 1 mL of medium appropriate for planned use. The cells are ready for subsequent experiments.
NOTE: However, with functional tests on fresh or frozen cells, a resting period in an incubator (typically overnight) is often recommended after lymphocyte isolation medium based isolation or cell thawing.
5. Cell culture
- After isolation or thawing culture cells overnight at 37 °C in 5% CO2/95% air.
- Resuspend the cells in 1 mL of RPMI medium supplemented with 10% FBS, penicilin 50 U/mL, streptomycin 50 U/mL. For further use there are many possibilities, treat cells as needed for the assays.
- For general storage use sterile 6 well cell culture plates and harvest cells after resting period. Transfer 1 mL of cell suspension with a 1 mL pipette into a well and add 4 mL of cell culture medium.
NOTE: The amount of PBMCs isolated varies greatly between individuals, one well with 5 mL of cell culture medium is sufficient for every 8 mL of blood isolated. However, when isolating PBMCs from buffy coats, the quantity of PBMCs is significantly higher than in whole blood samples and the cells should therefore be divided into several wells. - Let cells rest 24 h in a humidified atmosphere supplemented with 5% CO2 at 37 °C. This incubation is performed for freshly isolated cells, as well as for cryopreserved cells.
6. ATP assay
- Resuspend thawed PBMCs in 1 mL of RPMI medium supplemented with 10% FBS, penicillin 50 U/mL, streptomycin 50 U/mL.
- Take a sample and determine the cell count, then perform a live-dead discrimination with trypan blue. Take 10 µL from the resuspended cells and mix with 90 µL cell culture medium. Then take 10 µL and mix with 10 µL of trypan blue. Place the cells either in a cell counting chamber or an automatic cell counter and determine the number of live/dead cells.
- Plate 100 µL of cells at a density of 1 x 105 cells/100 µL per well in a 96-well white polystyrene plate.
- Let cells rest for 24 h in a humidified atmosphere supplemented with 5% CO2 at 37 °C.
- Determine the ATP concentrations with an ATP assay.
- Use light emission that occurs when ATP is combined with luciferin. The emitted light can be evaluated with a plate reader. Remove the plates for the incubator to cool down to room temperature for 15 min. Lyse the cells and leave them for 5 min. Then apply monitoring reagent to the cells and measure according to the manufacturer's instructions. An internal standard is used to determine the ATP level.
7. High-resolution respirometry
- Turn ON the high-resolution oxygraph and let it warm up for 30 min.
- Treat cells according to a protocol described in Figure 1. Prepare all stocks needed as in Table 1.
- Pipette 2.1 mL of respiration buffer (Table 1) into both high-resolution oxygraph chambers and stir the buffer continuously using a magnetic stirring bar present in the chambers (750 rpm) at 37 °C for 30 min until a stable oxygen flux signal of the polarographic oxygen sensor is obtained.
NOTE: In the chambers of the oxygraph, the oxygen consumption in real time (flux) and the oxygen saturation of the chamber are measured with the aid of polarographic oxygen electrodes. Background calibration must be performed to avoid background noise and to ensure reliable results. - Perform an air calibration of the polarographic oxygen sensor according to the manufacturer's protocols23.
- Resuspend isolated PBMCs in 1 mL of mitochondrial respiration medium (MIR05 the composition is shown in Table 1) and dilute to 8 x 106 cells/mL.
- After air calibration, aspirate the respiration medium from the oxygraph chamber and add 2.1 mL of cell suspension in each camber of the respirometer. If needed during the measurement reoxygenate the chambers (see Open at point h in Figure 2), the oxygen saturation of the chambers should not fall below 100 µM.
- Close the chambers by inserting the stoppers, the chambers are designed to hold 2.0 mL volume. Aspirate the emerging cell suspension.
- Continuously mix the cell suspension at 37 °C with a magnetic stirrer (750 rpm) located in the chamber. Wait for about 20 min until a stable signal is obtained. Determine endogenous respiration ((a) in Figure 2).
- To determine the different complex activities of the respiratory chain, inject the substrates and inhibitors for mitochondrial respiration through the titanium injection ports of the stoppers. Use the following final concentration in the chamber.
- To disrupt cell membranes, add 5 µL of 8.1 mM digitonin, through the titanium injection port of the chamber stopper, to remove naive substrates (b) in Figure 2 while the mitochondrial membranes remain intact.
- Add substrates 2 M glutamate and 800 mM malate through the injection port of the chamber stopper and record respiration until a stable signal is achieved. The signal stabilizes after 2-4 min.
NOTE: It is also possible to use additional substrates like pyruvate. - Add 8 µL of 500 mM adenosine diphosphate (ADP) through the injection port of the chamber stopper and record respiration until a stable signal is achieved. The signal stabilizes after 2-4 min (d) in Figure 2.
- Add 20 µL of 1 M succinate through the injection port of the chamber stopper and record respiration until a stable signal is achieved. The signal stabilizes after 2-4 min (e) in Figure 2.
- Titrate 1 M carbonyl cyanide p-trifluoromethoxy phenylhydrazone (FCCP) stepwise at 0.5 µL up to the point where no further increase occurs. Wait 2-4 min until the signal stabilizes (f) in Figure 2. When there is no further increase in respiration continue with the next step.
CAUTION: Be careful when handling FCCP as it has health risks for humans. - Add 5 µL of 0.1 mM rotenone through the injection port of the chamber stopper and record respiration until a stable signal is achieved. The signal stabilizes after 2-4 min (g) in Figure 2.
CAUTION: Be careful when handling rotenone as it has health risks for humans. - Add 1 µL of 4 mg/mL oligomycin through the injection port of the chamber stopper and record respiration until a stable signal is achieved. The signal stabilizes after 2-4 min (h) in Figure 2.
CAUTION: Be careful when handling oligomycin as it is a poison that poses health risks to humans. - Add 1 µL of 5 mM antimycin A through the injection port of the chamber stopper and record respiration until a stable signal is achieved. The signal stabilizes after 2-4 min (i) in Figure 2.
CAUTION: Be careful when handling antimycin A as it is a poison that poses health risks to humans. - When evaluating the run to exclude the oxygen consumption of enzymes not involved in oxidative phosphorylation, subtract the antimycin A values from all other measured values.
- Add 200 mM N,N,N',N'-tetramethyl-p-phenylenediamine dihydrochloride (TMPD; electron donator) and 800 mM ascorbate to keep TMPD in a reduced state. Inject the substrates through the injection port of the chamber stopper and record respiration until a stable signal is achieved. The signal stabilizes after 2-4 min (j) in Figure 2.
- TMPD is subject to autooxidation, so subtract the resulting oxygen consumption from the measured value at Complex IV.
- Add NaN3 ≥ 100 mM through the injection port of the chamber stopper and record respiration until a stable signal is achieved. The signal stabilizes after 2-4 min to inhibit complex IV activity, only TMPD autooxidation remains.
CAUTION: Be careful when handling sodium azide as it is a poison that poses health risks to humans.
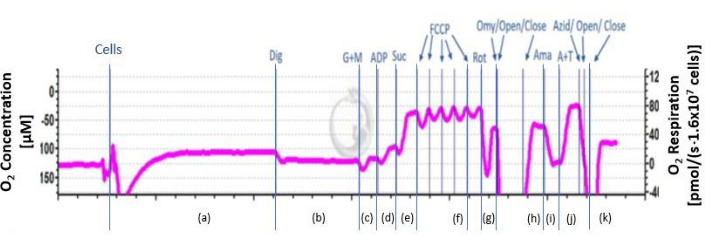
Figure 2: Schematic course of the O2 flux. The schematic course of the oxygen flux is shown. The curve is divided into the different phases after the addition of inhibitors and substrates from a-k. a: endogenous respiration; b: permeabilized cells; c: uncoupled complex I respiration; d: coupled complex I respiration; e: OXPHOS ; f: maximal uncoupled activity of CI and CII ; g: uncoupled respiration of complex II; h: leak respiration; i: residual respiration; j: CIV(U) uncoupled respiration and autooxidation of TMPD; k: autooxidation of TMPD. Please click here to view a larger version of this figure.
8. Citrate synthase activity
- Measure citrate synthase activity as a separate parameter and use it to normalize the measurements of the high-resolution oxygraph.
- Resuspend isolated PBMC in 1 mL of mitochondrial respiration medium (MIR05) and dilute to 8 x 106 cells/mL.
- Freeze in liquid nitrogen and store at −80 °C until experiments are conducted or to measure fresh cells.
- Prepare all solutions needed: 0.1 M triethanolamine HCl buffer pH 8.0, 1.0 M Tris-HCl buffer pH=8.1, 10% Triton X-100, 10 mM Oxalacetate in 0.1 M triethanolamine HCl buffer pH 8.0, 1.01 mM DTNB in 1.0 M Tris-HCl buffer pH=8.1, Acetyl-CoA 12.2 mM in double distilled H2O.
- Prepare reaction medium (Table 1) containing 5,5'-dithio-bis-(2-nitrobezoic acid) (DTNB; 0.1 mM), acetyl coenzyme A (0.31 mM), EDTA (50 µM), triethanolamine HCl (5 mM) and Tris HCl (0.1 M).
- Prepare starting reagent (Table 1) with 0.5 mM oxaloacetate dissolved in double distilled H2O.
- Thaw the samples on ice as citrate synthase is unstable if thawed too quickly.
- Add 40 µL of the samples to a 96-well plate on ice before adding 110 µL of reaction medium with a multipipette.
- Warm reaction medium and sample in an incubator to 30 °C for 5 min. Warm starting reagent in a water bath to 30 °C for 5 min.
- Add 50 µL of the starting reagent with a multipipette to each well. Measure absorbance at 30 °C at a wavelength of 412 nm for 20 min via plate reader.
Cell viability and number
To achieve successful isolation and cryopreservation, cell count and viability should be as high as possible. Before and after cryopreservation, the cells are counted, and their viability is determined to ensure the health and quality of the cells. Figure 3 is a representative illustration of PBMCs before and after cryopreservation, cell count and viability hardly differ. This indicates successful isolation and preservation of PBMCs.
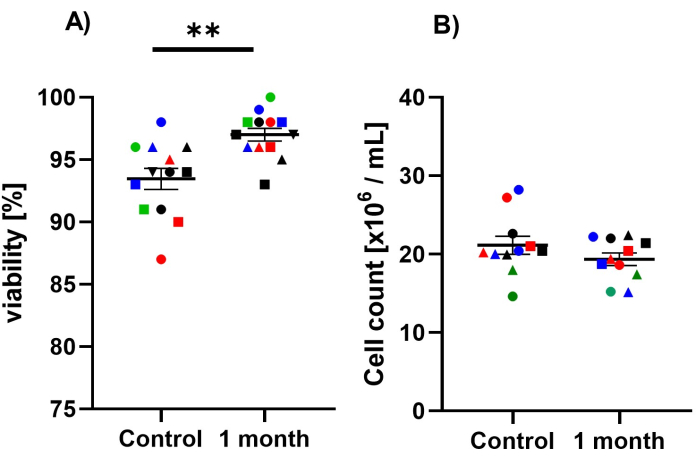
Figure 3: Effect of cryopreservation on cell number and viability. The test groups are divided into the control group with freshly isolated PBMCs and PBMCs after 1 month cryopreservation. The counting of the cells and the determination of their viability was carried out with an automated cell counter. Trypan blue was used to determine the viability. Each measurement was carried out as a triplicate of the mean value was calculated from the results of the individual measurements. (A) Determination of cell viability of PBMCs after fresh isolation and after 1 month cryopreservation. Values are given as mean ± SEM. Significances were determined with a paired t-test. (B) Determination of cell number of PBMCs after fresh isolation and after 1 month cryopreservation. Values are given as means ± SEM. Significances were determined with a paired t-test. The same shape and color show the same sample before and after one month of cryopreservation. Please click here to view a larger version of this figure.
ATP represents the major source of energy for eukaryotic cells. ATP is determined via a luminescence assay system. If cryopreservation is successful ATP between freshly isolated cells and cryopreserved cells should not differ. Figure 4 is a representative illustration of PBMCs before and after cryopreservation; ATP-levels are similar. This indicates successful isolation and preservation of PBMCs.
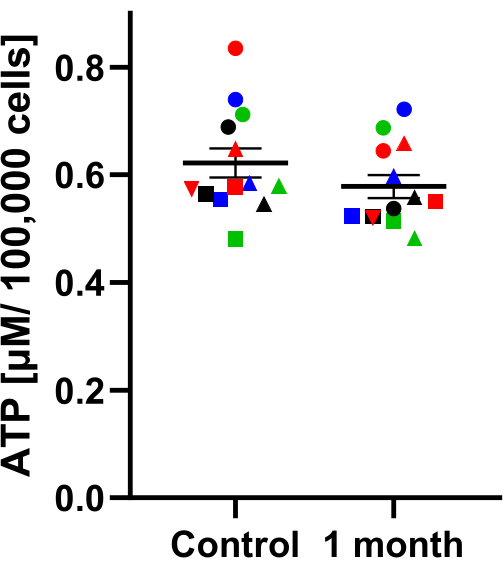
Figure 4: Comparison of ATP concentration of different cryopreservation periods. ATP concentration (µM / 100,000 cells) in PBMCs. The test groups are divided into the control group with freshly isolated PBMCs and PBMCs after 1 month cryopreservation. Samples were collected at the same time and then either cryo-stored or measured after isolation. A paired t-test was performed to test significant differences; the result showed that the differences were not significant. The values are given as mean values ± SEM (N = 13). The same shape and color show the same sample before and after one month of cryopreservation. Please click here to view a larger version of this figure.
Citrate synthase (CS) is a key enzyme of the citrate-cycle, located in mitochondria. Therefore, CS activity represents a robust marker for mitochondrial mass. CS activity is determined on the basis of conversion from DTNB into TNB. Figure 5 shows that before and after cryopreservation, CS values do not differ in PBMCs. Again, this indicates a successful isolation and preservation of PBMCs.
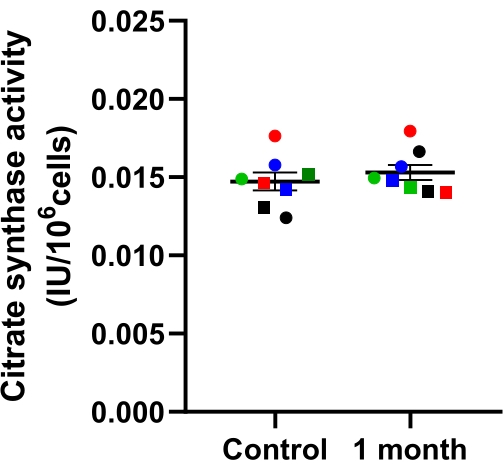
Figure 5: Effect of cryopreservation on citrate synthase activity. Citrate synthase activity in PBMCs after 1 month cryopreservation compared to the freshly measured control. Samples were collected at the same time and then either cryo-stored or measured after isolation. Values are expressed as mean ± SEM (N = 8). Significances were determined with a paired t-test. The same shape and color show the same sample before and after one month of cryopreservation. Please click here to view a larger version of this figure.
The bioenergetic profiles of PBMCs can be determined using polarographic oxygen sensors, e.g., in high-resolution oxygraph. Cellular oxygen consumption is measured in a closed-chamber system with very high resolution and sensitivity in biological samples, e.g., intact and permeabilized cells, tissues, or isolated mitochondria. The high-resolution oxygraph device is equipped with two chambers and uses polarographic oxygen sensors to measure oxygen concentration and calculate oxygen consumption within each chamber. Oxygen consumption rates are calculated using software and expressed as picomoles per s per number of cells24. Within each oxygraph chamber polarographic oxygen electrodes measure the oxygen concentration and calculate oxygen consumption (flux) within each chamber. Oxygen consumption and concentration in the chamber is displayed in real time (Figure 6). With the addition of specific inhibitors and substrates, the individual complexes of the respiratory chain are targeted in order to measure their activity. After the assay, the data is analyzed with the software. The flux values were normalized to citrate synthase activity. The oxygraphy provided lower values for complex IV due to partially oxidized TMPD. In a series of tests, we found that the TMPD used deviated by a factor of 1.8. This factor was used to normalize the values in Figure 6.
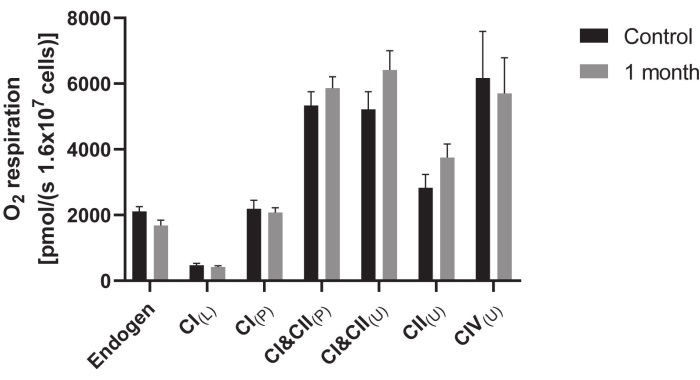
Figure 6: Mitochondrial respiration in PBMCS after cryopreservation compared to freshly measured control. A solution containing 1.6 x 107 cells/mL was used to measure the oxygen consumption of cells in an oxygraph. Respiration was measured 1 month after cryopreservation. To investigate the activity of the complexes in the respiratory chain several inhibitors, substrates and uncouplers were added. The addition of a substance was done as follows: CI(L) = leak respiration of complex I; CI(P) = coupled respiration of complex I; CI&CII(P) = physiological respiration; CI&CII(U) = uncoupled respiration displaying maximum activity of complexes I and II; CII(U) = uncoupled respiration of complex II; CII(L) = leak respiration of complex II; CIV(U) = uncoupled respiration. A factor of 1.8 was used to normalize the values. This factor was determined experimentally for the present setup. Data are displayed as the means ± SEM (N = 10). Statistical significance was tested via Student's t-test (*p < 0.05). Please click here to view a larger version of this figure.
Endogenous respiration is determined by adding 2 mL of cell suspension into the chambers. The flux with endogenous substrates is measured. To further distinguish between complexes digitonin is added. Digitonin permeabilizes the plasma membrane while the mitochondrial membranes remain intact. To compensate for proton leak through the membrane, substrates glutamate and malate are added. The respiration rates at this point illustrate complex I-driven respiration in the absence of coupled respiration. To detect the oxidative phosphorylation (OXPHOS) capacity of complex I ADP is added, the respiration is now in a coupled state. The coupled respiration of CI and CII is achieved by addition of succinate. Now the respiratory chain works at maximal capacity. With the titration of carbonyl cyanide p-trifluoromethoxy phenylhydrazone (FCCP) the electron transport chain (ETC) is uncoupled. With this uncoupling the maximal uncoupled activity of CI and CII is determined. To differentiate between CI and CII the complex I specific inhibitor rotenone is added. By the addition of oligomycin, the leak respiration of CII is determined. To exclude oxygen consumption by sources not involved in oxidative phosphorylation, the antibiotic antimycin A is added and this residual oxygen consumption is subtracted from all readings obtained in the experiment. The electron donor N,N,N',N'-tetramethyl-p-phenylenediamine dihydrochloride (TMPD; 0.5 mM) is an artificial substrate for CIV. To keep TMPD in a reduced state, ascorbate is used. Ascorbate and TMPD are used to measure the maximum uncoupled respiration of CIV. Since TMPD is subject to auto-oxidation, NaN3 is added after stabilization of the flux to inhibit CIV. The remaining oxygen consumption is subtracted from the raw CIV values. Figure 2 shows a typical measurement curve.
The parameters shown show the success of the technique. The technique was developed to perform bioenergetic measurements after cryopreservation. The results shown compare the cells before and after the measurement, as no statistically significant differences can be seen, it can be assumed that this method of preservation is suitable for storage over 1 month.
Table 1: Solutions, buffers, and consumables. Please click here to download this Table.
