The yields of mitochondria obtained following the above-described protocols vary depending on several factors such as the cell line or tissue type, the nature of the samples (i.e., if fresh or frozen tissues are used), or the efficiency of the homogenization process. Expected yields of mitochondria from different cell lines and tissues are collected in Table 2. Once the mitochondrial fractions have been obtained, the next step is the analysis of respiratory SCs pattern, which is performed after the crude mitochondrial sample solubilization and electrophoretic separation by BN-PAGE followed by IGA-analysis or WB immunodetection. Figure 1 shows the aspect of unstained gel lanes just after the run and the pattern of bands after standard Coomassie staining of a cultured cell line and a mouse liver mitochondrial sample.
In Figure 2A,B, clear differences between the SC assembly patterns in human and mouse cells after using IGA assays can be observed. Thus, free complex I is observed in mouse cells whereas it is not detectable in human mitochondria. Complex IV-IGA pattern is very similar in both human cell lines (Figure 2B) while presenting differences between the two mouse cell lines analyzed in Figure 2A. These differences are due to the fact that BL6 cells express a mutant variant of SCAF18.
The SC patterns, obtained with the protocol for mitochondrial isolation described here to work with small samples, are maintained with respect to the "conventional" protocols used for larger samples. This can be seen in Figure 2C, by comparing lanes 1 and 6 (obtained after mitochondrial isolation using a conventional protocol from 3 g of liver and 1.1 g of BAT) with, respectively, lanes 2 and 5 (obtained using the protocols here presented from around 0.1 g of both tissues). As proposed in the plasticity model, the relative proportion of respiratory complexes and SCs varies depending on the cell type and the metabolic state. As shown in Figure 2C,D, the different tissues show different SC assembly patterns. Thus, brain mitochondria show very low levels of free complex I and a higher proportion of SCs when compared with the other tissues, and BAT is characterized by its low levels of CV (Figure 2A) and high amounts of SC III + IV (Figure 2D). Heart mitochondria present high levels of SCs and CV. CV assembly patterns are very similar between the cultured cell line and a mouse liver mitochondrial sample (Figure 2E).
SC assembly can also be analyzed by WB as shown in Figure 3 for two samples obtained from spleen control and tumor cells. In this case, 30-50 µg of mitochondrial protein are usually enough to detect the different complexes and SCs with the specific antibodies. Figure 4 shows some of the main pitfalls that can happen during the application of these protocols. Wrong gel gradients can produce abnormal and undesired migration patterns as in Figure 4A, lane 1, where the SCs remain close to the well in the upper part of the lane due to a higher-than-normal acrylamide concentration in the gel. Incubation of the samples, even for relatively short times, at high temperatures (here 37 °C and 40 °C for 10 min) causes degradation of SCs and CI (Figure 4A lanes 2-4). Likewise, the detergent-to-protein ratio is critical for good results and adequate gel resolution. Below a ratio of 2 g of digitonin/g of protein, the band resolution is progressively reduced and at the lower concentration (0.5 g/g) only a smear in the area corresponding to SCs is visible (Figure 4B).
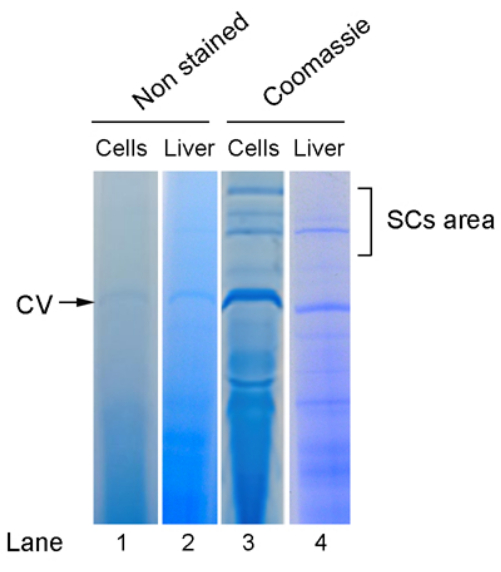
Figure 1: Pattern of mitochondrial complexes and supercomplexes after BNGE separation of digitonin-permeabilized mitochondria from a culture cell line (L929Balbc) and mouse liver. The lanes on the left (1 and 2) represent the aspect of the unstained gel just after the run, where only CV is detectable, while the lanes on the right (3 and 4) show the pattern of bands after Coomassie staining. Abbreviations: BNGE = Blue Native gel electrophoresis; CV = complex V; SCs = supercomplexes. Please click here to view a larger version of this figure.
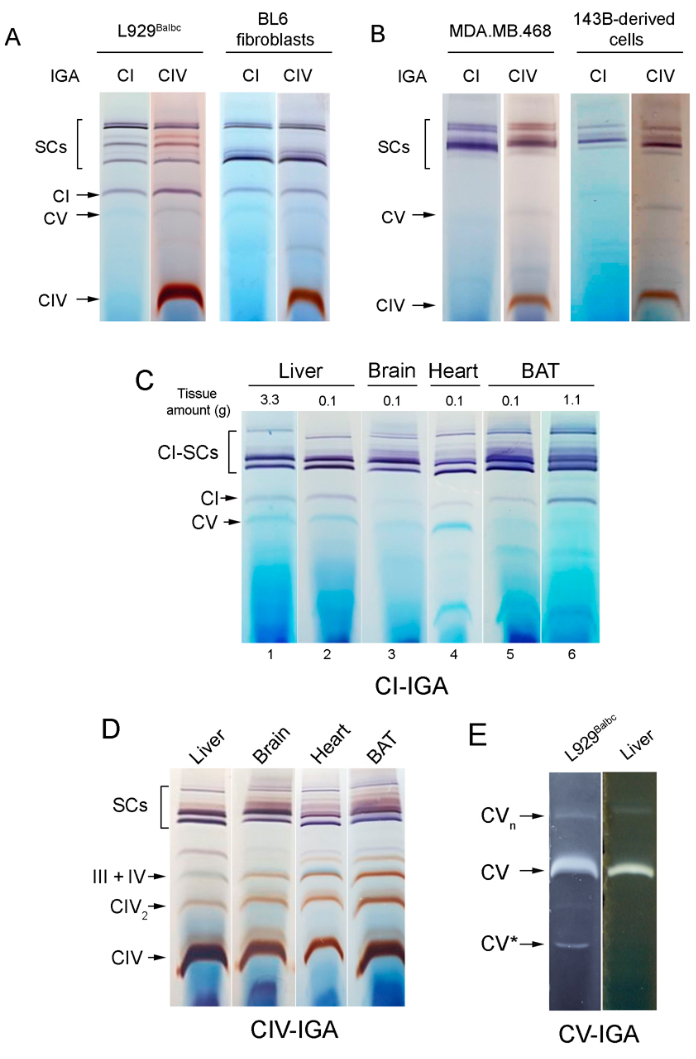
Figure 2: IGA-analysis of mitochondria isolated from different cell lines and mouse tissues. (A,B) In-gel activity of the indicated complexes in mitochondria isolated from (A) mouse or (B) human cell lines. L929Balbc is a transmitochondrial cell line generated in our laboratory by transferring mitochondria from Balb/cJ mouse platelets to ρ°L929neo cells as described previously40,41. BL6 fibroblasts are an immortalized cell line generated in our lab from an ear biopsy of a BL6 mouse. (C) CI-IGA patterns from mouse mitochondria isolated using either a conventional method starting with 3 g of liver (lane 1) and with 1.1 g of BAT (lane 6) or the protocol presented here and starting with around 0.1 g of tissue in all cases (lane 2, liver; lane 3, heart; lane 4, brain, and lane 5 BAT). (D) CIV-IGA patterns (performed after CI-IGA shown in panel 2C, lanes 2-5) in mouse liver, heart, brain, and BAT mitochondria. (E) CV-IGA activity analyzed in mouse cultured cells and liver. Around 60-75 µg of mitochondrial protein per lane were loaded. Abbreviations: IGA = in-gel activity; SCs = supercomplexes; CI = complex I; BAT = brown adipose tissue. Please click here to view a larger version of this figure.
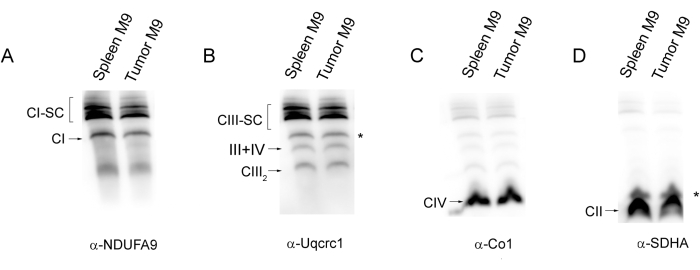
Figure 3: WB immunodetection of the different mitochondrial complexes and supercomplexes in spleen samples. After Blue Native PAGE separation of mitochondrial complexes and SCs obtained from spleen control (Spleen M9) and tumor (Tumor M9) samples, they were transferred to a PVDF membrane and sequentially hybridized with antibodies that recognize the indicated CI, CIII, CIV, and CII subunits (panels A–D, respectively). Around 50 µg of mitochondrial protein were loaded per lane. Asterisks indicate the signal remaining from a previous western blot. Abbreviations: SC = supercomplex; CI = complex I; CIII = complex III; CIV = complex IV; CII = complex II. Please click here to view a larger version of this figure.
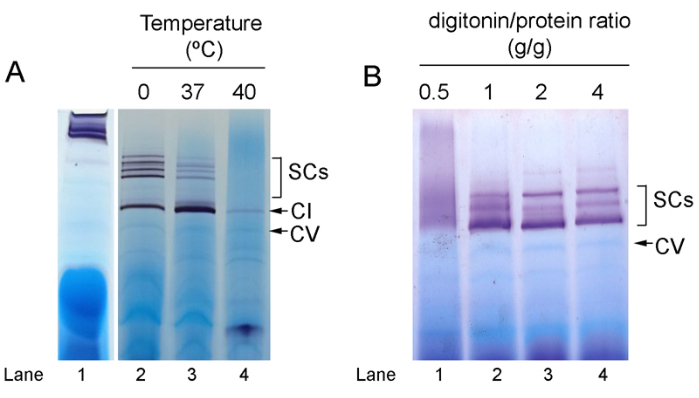
Figure 4: Pitfalls in SC analysis by Blue-Native PAGE. CI IGA-analysis of mitochondria isolated from (A) fresh mouse liver and (B) frozen rat liver under different solubilization conditions. (A) Lane 1, wrong gel gradient concentration (8-13% instead of the normal 3-13%). Lanes 2-4, samples incubated for 10 min on ice or at 37 °C and 40 °C before loading, respectively. (B) Pattern after IGA for CI of mitochondria obtained from frozen rat liver using different digitonin/protein ratios for solubilization. Abbreviations: SC = supercomplex; PAGE = polyacrylamide gel electrophoresis; CI = complex I; IGA = in-gel activity. Please click here to view a larger version of this figure.
| Medium | Composition | ||
| Hypotonic buffer | 10 mM MOPS, 83 mM sucrose, pH 7.2 | ||
| Hypertonic buffer | 30 mM MOPS, 250 mM sucrose, pH 7.2 | ||
| Homogenization Buffer A | 10 mM Tris, 1 mM EDTA, 0.32 M sucrose, pH 7.4 | ||
| Homogenization Buffer AT | 225 mM mannitol, 1 mM EGTA, 75 mM sucrose, 0.01% BSA pH 7.4 | ||
| Homogenization Buffer AT2 | 225 mM mannitol, 1 mM EGTA, 75 mM sucrose, 0.02% BSA pH 7.4 | ||
| BN sample buffer | 50 mM NaCl, 50 mM Imidazole, 2 mM Aminocaproic acid, 1 mM EDTA pH 7.0 | ||
| Digitonin solution | 10% digitonin in 50 mM NaCl, 50 mM Imidazole, 5 mM Aminocaproic, 4 mM PMSF | ||
| BN loading buffer | 5% Coomassie Blue G in 0.75 M aminocaproic acid | ||
| BN cathode buffer A | 50 mM tricine, 15 mM Bis-Tris pH =7.0 (4 °C), 0.02% G-250 (Coomassie brilliant blue G-250) | ||
| BN cathode buffer B | 50 mM tricine, 15 mM Bis-Tris pH=7.0 (4 °C), 0.002% G-250 | ||
| BN anode buffer | 50 mM Bis-Tris, pH=7.0 | ||
| Complex I IGA substrate | 5 mM Tris-HCl pH 7.4, 0.1 mg/mL NADH, 2.5 mg/mL Nitroblue Tetrazolium (NBT) | ||
| Complex II IGA substrate | 50 mM Potassium phosphate buffer pH 7.4, 20 mM sodium succinate, 0.2 mM phenazine methosulfate (PMS), 2.5 mg/mL NBT | ||
| Complex III IGA substrate | 50 mM Potassium phosphate buffer pH 7.2, 0.05% diaminobenzidine (DAB) | ||
| Complex IV IGA substrate | Same solution as CIII plus 50 μM cytochrome c | ||
| Complex V IGA substrate | 35 mM Tris, 270 mM glycine pH 8.3, 14 mM MgSO4, 0.2% Pb(NO3)2, 8 mM ATP | ||
| Staining solution | Coomassie dye R-250 at 0.25% in 40% methanol and 10% Acetic acid | ||
| Destaining/fixing solution | 40% Methanol, 10% Acetic Acid | ||
| Transfer buffer | 48 mM Tris, 39 mM Glycine, 20% methanol | ||
Table 1: Buffers and media composition.
| Cell type/tissue | Starting amount | Expected yield (µg mitochondrial protein) | Number of lanes (50-60 µg/lane) |
| Cultured cells | |||
| MDA-MB-468 | 100 µL vpc | 300-400 µg | 6-9 |
| 143B-derived | 100 µL vpc | 200-350 µg | 4-7 |
| Mouse Fibroblasts | 100 µL vpc | 150-200 µg | 3-5 |
| Heart | 100 mg | 400-500 µg | 10-12* |
| Skeletal muscle | 100 mg | 300-400 µg | 8-10* |
| Liver | 100 mg | 400-500 µg | 8-10 |
| Spleen | 50 mg | 150-250 µg | 3-5 |
| BAT | 100 mg | 200-300 µg | 4-6 |
| Brain | 100 mg | 250-300 µg | 5-8 |
Table 2: Expected yields of mitochondria from different cell types and mouse tissues. The mitochondrial fractions obtained from heart or skeletal muscle are purer than those obtained from other tissues. Thus, smaller amounts can be loaded in the gels (*).
