Determine the distribution of BEV-enriched fractions
To determine the distribution of bacterial extracellular vesicles (BEVs)-enriched fractions, a blank control was established to measure the absorbance values at OD 340 nm, and the density of each fraction was calculated based on the measurements and iodixanol guidelines (Step 8.1). Table 2 presents the density results, demonstrating that fractions F4 to F9 exhibited densities within the range typically associated with extracellular vesicles. This finding suggested that the majority of BEVs were isolated in these fractions, leading to the definition of F4-F9 as the rough range for BEVs. Transmission electron microscopy (TEM) was employed to characterize the morphological features of fecal BEVs isolated as shown in Figure 1 in fractions F4-F9. The TEM images (Figure 2) revealed the presence of classical cup-shaped structures, which are characteristic of BEVs. Nanoparticle tracking analysis (NTA) and bicinchoninic acid assay (BCA) were conducted to determine the particle number and protein concentration, respectively, allowing for further evaluation of recovery rates and purity. Figure 3 displays the protein and particle concentrations for each fraction, indicating that F6 and F7 exhibited the highest concentrations, followed by F5 and F8. Subsequently, Coomassie brilliant blue staining (CBBS) and western blotting (WB) were performed to provide an overall protein analysis. The results of CBBS were consistent with the protein concentration findings, with F6 and F7 displaying the most intense bands (Figure 4). To confirm the distribution of BEVs, extracellular vesicles derived from Escherichia coli were employed as a positive control (defined in Step 8.5.13). The isolation and purification procedures followed the steps described in the Table of Materials and the aforementioned protocol sections (Steps 3, 4). WB analysis further confirmed the presence of outer membrane protein A (OmpA), a major component of the outer membrane of bacteria and a specific marker for BEVs, in fractions F6-F8. Based on these results, fractions F6-F8 were defined as BEV-enriched fractions suitable for subsequent experiments and analysis.
Verification of the range of presentation of the interference components
Eukaryotic extracellular vesicles (EEVs), which possess a similar size and density as BEVs, were identified as a major interference in the analysis of BEVs. To address this, three EEV proteins were probed in fractions 5-8 from both the Top-down and Bottom-up modes, with centrifugation times of 7 h and 15 h. CD63, an EEV-associated transmembrane protein, was predominantly detected in F5, while the BEV marker LPS was enriched in fractions 6-7. This pattern was observed in both modes, although the Bottom-up mode exhibited slight banding of CD63-EEVs in F7. However, other EEV markers, CD9 and TSG-101, did not show visible signals in these fractions regardless of the modes or centrifugation times. In an independent experimental undertaking, antibodies targeting Syntenin and Integrin β1 were employed to ascertain the enrichment of distinct protein markers inherent to EEVs across fractions 5-8. Syntenin was prominently detected within F5, particularly under the ambit of the Top-down approach, while the presence of Integrin β1 was discerned within F6 (Figure 5). To further assess the presence of interfering particles, Dil-labeled low-density lipoprotein (Dil-LDL) was introduced into the density gradient system, and fluorescence intensity was measured across all fractions. In the Top-down mode, fractions 1-4 exhibited relatively higher fluorescence values, while in the Bottom-up mode, it was F1-F6 that displayed higher intensities (Figure 6).
Assess the recovery rates and purity of two DGC modes from concentration, particle size, and markers of BEVs
After identifying F6-F8 fractions as BEV-enriched fractions, the recovery rates and purity of the two density gradient centrifugation (DGC) modes were evaluated. The particle sizes of BEV-enriched fractions obtained from the Top-down mode, Bottom-up mode, and ultracentrifugation (UC) alone were compared. The observed particle sizes in both centrifugation modes ranged from 90 to 300 nm, aligning with the defined diameter of BEVs (Figure 7). Next, the particle and protein recovery rates of the two modes were calculated based on the particle and protein concentrations before and after DGC (Table 3 and Figure 8). Notably, the Bottom-up mode exhibited higher recovery rates compared to the other mode. In a separate experimental group, protein recovery in the two modes was evaluated at different centrifugation times of 7 h and 15 h. Importantly, there was no statistically significant difference in protein content within F6-F8 fractions after DGC between the two centrifugation times, regardless of the gradient mode used (Figure 9). The particle/protein ratio was calculated to provide a rough assessment of the purity in the two modes. The data indicated no significant difference in purity between the modes. Additionally, Coomassie brilliant blue staining (CBBS) and western blotting (WB) were performed to further compare the purity, focusing on three BEV markers (LPS, OmpA, and LTA), one EEV marker (CD63), and two contaminating markers (Calnexin, Flagellin). The results demonstrated a partial reduction of interfering substances after DGC, with LPS and OmpA showing a more prominent presence in both DGC modes compared to UC alone (Figure 10).
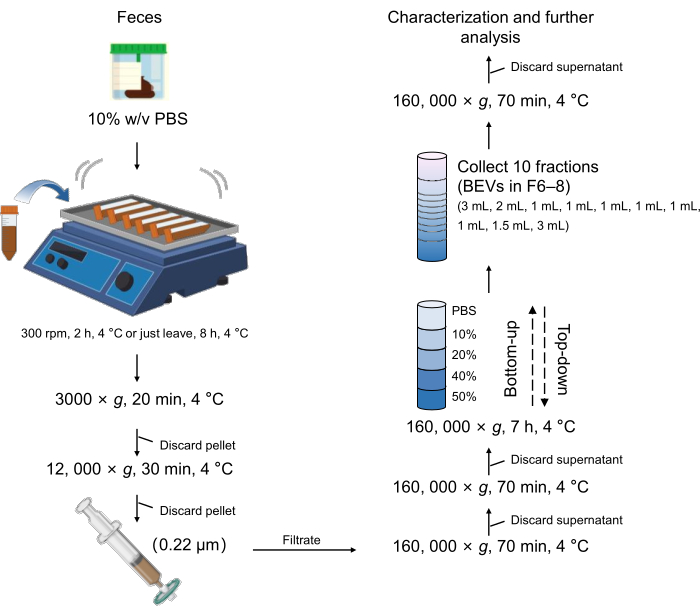
Figure 1: Schematic workflow of BEVs isolation and purification. Abbreviations: BEVs = bacterial extracellular vesicles. Please click here to view a larger version of this figure.
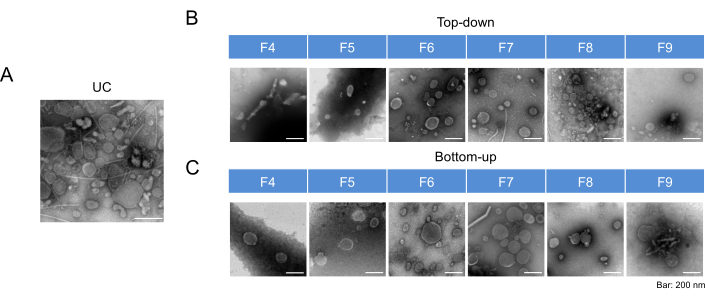
Figure 2: Representative TEM images of BEV fractions. (A) TEM images of BEVs isolated after UC. (B) TEM images of BEVs isolated after DGC using the Top-down mode. (C) TEM images of BEVs isolated after DGC using the Bottom-up mode. All images were presented at a consistent scale of 200 nm. Abbreviations: TEM = transmission electron microscope; BEVs = bacterial extracellular vesicles; UC = ultracentrifugation; DGC = density gradient centrifugation. Please click here to view a larger version of this figure.
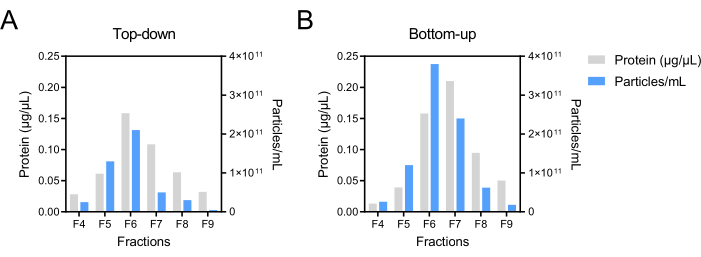
Figure 3: Protein and particle concentration of BEV fractions after Top-down and Bottom-up DGC modes. The protein concentration of BEV fractions was determined using BCA and is represented by the grey bars. The particle concentration was measured using NTA and is represented by the blue bars. Abbreviations: BEV = bacterial extracellular vesicle; DGC = density gradient centrifugation; BCA = bicinchoninic acid assay; NTA = nanoparticle tracking analysis. Please click here to view a larger version of this figure.
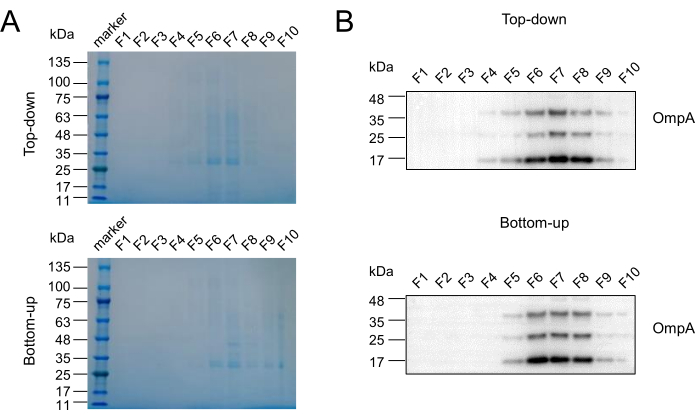
Figure 4: Determination of BEV-enriched fractions. (A) CBBS was performed to determine the fractions enriched with fecal BEVs in both Top-down and Bottom-up modes. (B) EVs derived from Escherichia coli were isolated, purified, and utilized as a positive control in WB to confirm the presence and distribution of BEVs. OmpA: Outer membrane protein A, a BEV marker. Abbreviations: BEV = bacterial extracellular vesicle; CBBS = coomassie brilliant blue staining; EVs = extracellular vesicles; WB = western blotting. Please click here to view a larger version of this figure.
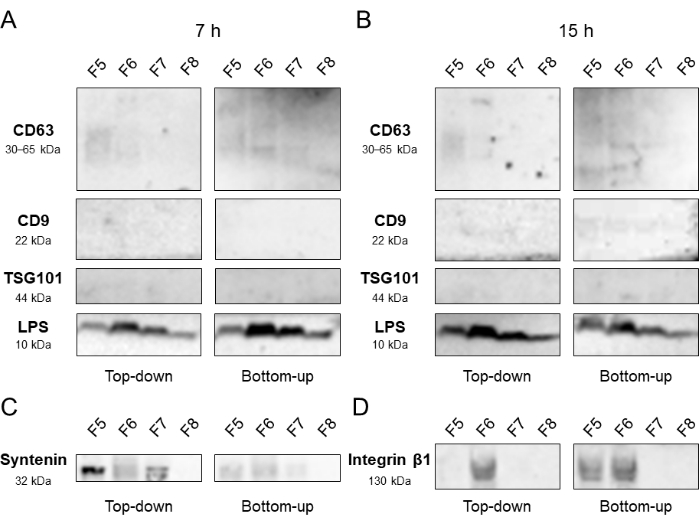
Figure 5: Confirmation of EEV distribution. Western blot analysis of fractions 5-8 obtained from the Top-down and Bottom-up DGC modes with different density gradient ultracentrifugation durations, 7 h (A) and 15 h (B). An independent trial focusing on F5 to F8 resulting from a 7-h centrifugation period was executed, as demonstrated in (C) and (D). Selected markers included those associated with BEVs (LPS) and EEVs (CD63, CD9, TSG-101, Syntenin, Integrin β1). Abbreviations: DGC = density gradient centrifugation; BEV = bacterial extracellular vesicle; EEV = eukaryotic extracellular vesicle. Please click here to view a larger version of this figure.
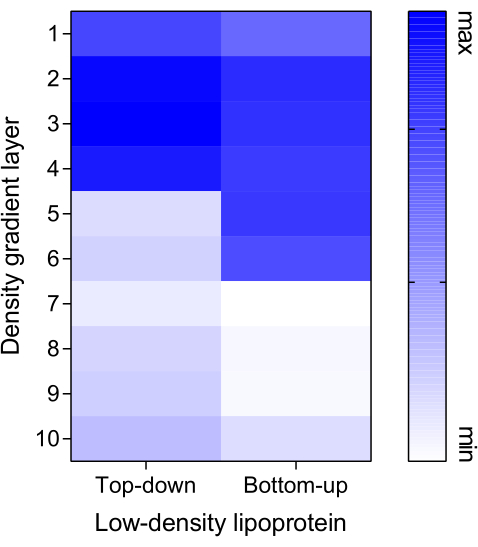
Figure 6: Distribution of low-density lipoprotein. Dil-labeled low-density lipoprotein (Dil-LDL), considered a contamination marker, was added at a concentration of 10 µg to the gradient system in both the Top-down and Bottom-up modes. Dil-LDL replaced the PBS/BEV solution (Step 6.1.1 and Step 6.2.2) to establish a detection model. The fluorescence intensity of each fraction was measured using a microplate detector with an excitation/emission wavelength of 549/565 nm. Abbreviation: BEV = bacterial extracellular vesicle. Please click here to view a larger version of this figure.
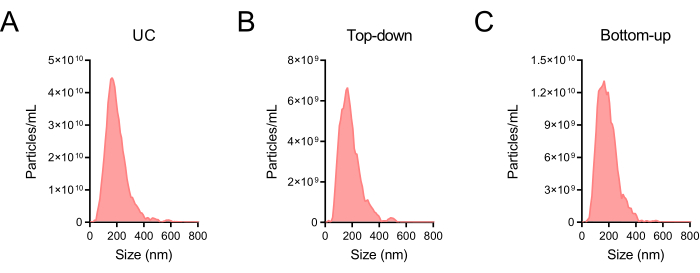
Figure 7: Particle sizes of BEV-enriched fractions. (A) Particle size distribution of crude BEVs obtained after UC. (B) Particle size distribution of BEV-enriched fractions (F6-F8) obtained using the Top-down density gradient centrifugation (DGC) mode. (C) Particle size distribution of BEV-enriched fractions (F6-F8) was obtained using the Bottom-up DGC mode. NTA was undertaken to quantify the dimensions of the particles. The dilution factors employed for panels (A–C) were 10,000, 2,000, and 5,000, correspondingly, with the ensuing outcomes calibrated. Abbreviations: BEV = bacterial extracellular vesicle; UC = ultracentrifugation; DGC = density gradient centrifugation; NTA = nanoparticle tracking analysis. Please click here to view a larger version of this figure.
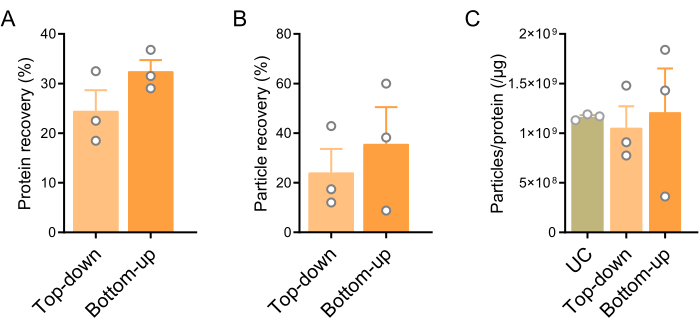
Figure 8: Protein and particle recovery rates of Top-down and Bottom-up DGC modes. (A) Protein recovery rates of the two modes calculated as the ratio of protein concentration after and before DGC (UC). (B) Particle recovery rates of the two modes are calculated by the ratio of particle concentration after and before DGC (UC). (C) Particle/protein ratios of UC, Top-down, and Bottom-up modes. Data are presented as mean ± SEM. Statistical analysis was performed using a two-tailed, unpaired t-test for panels A and B, and one-way analysis of variance (ANOVA) with Tukey post hoc test for panel C. No significant differences were observed. Abbreviations: DGC = density gradient centrifugation; UC = ultracentrifugation. Please click here to view a larger version of this figure.
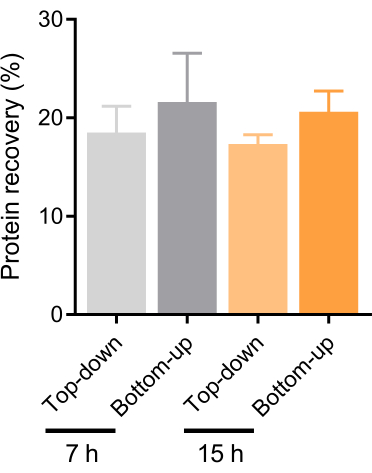
Figure 9: Comparison of protein recovery based on Top-down and Bottom-up DGC modes using different density-associated ultracentrifugation times. Protein recovery rates in fractions F6-F8 obtained from gradient ultracentrifugation for 7 h and 15 h were evaluated in independent experiments using both Top-down and Bottom-up modes. Data are presented as mean ± SEM. Statistical analysis was performed using two-way analysis of variance (ANOVA) with Fisher's least significant difference test. No significant differences were observed. Abbreviations: DGC = density gradient centrifugation. Please click here to view a larger version of this figure.
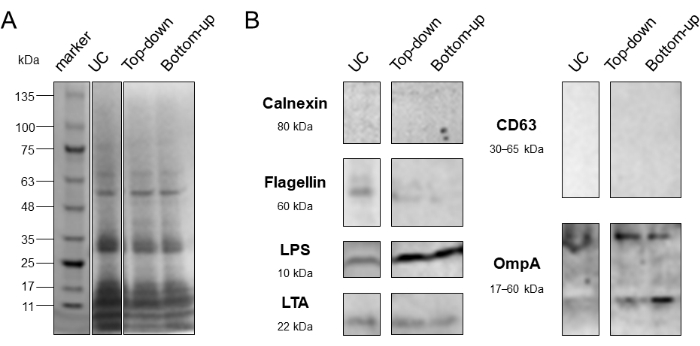
Figure 10: Purity assessment of Top-down and Bottom-up DGC modes. (A) CBBS was conducted to compare the protein distribution of UC, Top-down, and Bottom-up modes. (B) WB was conducted to assess the purity of UC, Top-down, and Bottom-up modes. Calnexin: endoplasmic reticulum-associated protein marker; Flagellin, contaminating protein marker from bacteria. CD63: transmembrane protein marker for eukaryotic extracellular vesicles; LTA: BEVs marker from gram-positive bacteria; LPS, OmpA: BEVs markers from gram-negative bacteria. Abbreviations: DGC = density gradient centrifugation; UC = ultracentrifugation; CBBS = Coomassie Brilliant Blue staining; WB = western blotting. Please click here to view a larger version of this figure.
| Iodixanol solution concentration (%) | 0.02 M HEPES buffer | 50% Iodixanol working solution |
| 40 | 1 | 4 |
| 20 | 3 | 2 |
| 10 | 4 | 1 |
Table 1: Ratios for preparing iodixanol density gradient buffers. The table provided the ratio of 0.02 M HEPES buffer and 50% iodixanol working solution to obtain a certain concentration of iodixanol solution.
| F1 | F2 | F3 | F4 | F5 | F6 | F7 | F8 | F9 | F10 | |
| Density (g/mL) | 1.023 | 1.038 | 1.049 | 1.062 | 1.074 | 1.098 | 1.144 | 1.185 | 1.239 | 1.293 |
Table 2: The density of the iodixanol-based density gradient centrifugation system. The table showed the density of each fraction in an iodixanol-based density gradient centrifugation system. Blank control: PBS-based gradient system.
| Top-down mode | Bottom-up mode | |
| Particle recovery rate (%) | 24.08 | 35.69 |
| Protein recovery rate (%) | 24.5 | 32.45 |
Table 3: The particle and protein recovery rates of two modes. The table showed the particle and protein recovery rates of the Top-down and Bottom-up modes (Figure 8).
