1. Step 1: Mouse Embryonic Stem (mES) Cell Culture (Timing: 3 days)
1. Plating primary embryonic mouse fibroblasts (PMEF)
- Coat four 100-mm tissue culture dishes with gelatin for PMEF adhesion. Add 8 ml of 0.1% gelatin solution (StemCell Technologies) to each dish and incubate for 30 min at room temperature.
- Remove excess gelatin from dishes. Do not rinse the dishes.
- Dilute one vial of Mitomycin C-inactivated PMEF (Millipore) that contains approx. 5 x 106 cells into 40 ml of PMEF media (see recipe in Table 1) and plate onto four 100-mm tissue culture dishes. The PMEF provide a feeder layer for mES cells.
- PMEF should be plated at least one day before adding mES cell cultures. PMEF can be used up to 1 week after plating.
2. Plating mES Cells
To monitor the efficiency of mES cell differentiation into motor neurons, we used HBG3, an ES cell line derived from a transgenic mouse expressing eGFP under the control of the motor neuron specific promoter Hb9.
- Defrost 1 vial of HBG3 mES cells (approx. 1 x 107, contributed by Dr. Douglas Kerr, Johns Hopkins University) in a 37 °C water bath. Add 5 ml of ES media (see recipe in Table 1) in a 15 ml tube and spin down cells at 800 rpm for 5 min.
- Aspirate supernatant and resuspend mES cells in 20 ml of ES media with freshly added Leukemia inhibitory factor (LIF) at a final concentration of 10 ng/ml. NOTE: LIF should be added on the day of use and is required to maintain the mES cells in an undifferentiated state.
- Remove 10 ml PMEF media from the PMEF in each dish and add 10 ml of the HBG3 ES cell suspension to each dish.
- 24 h after plating, mES cells form small colonies, similar in size to the colony indicated by the arrowhead in Figure 1B. 72 h after plating, colonies increase in size, many being approximately 100 μm in diameter (Figure 1B, indicated by arrows). These cells are ready for differentiation. Media should be replaced daily to maintain proliferation of cells.
2. Step 2: Neural Induction (Timing: 2 days)
To induce differentiation of mES cells into motor neurons, mES cells need to be separated from PMEF and cultivated in a suspension environment. Gelatin coated flasks are used to separate PMEF from mES cells.
- When mES colonies are ready for neural induction (defined as differentiation day 0), coat 2 T150 flasks with 0.1% gelatin (18 ml/flask) for 30 min at room temperature and then remove excess gelatin and wash with 1x PBS three times. NOTE: Each 100-mm dish culture will need one T150-gelatin coated flask to separate PMEF.
- Carefully remove media by aspiration, making sure not to dislodge loosely attached colonies. Add 10 ml PBS briefly and then remove by aspiration to complete wash step.
- To separate mES cells from PMEF, add 0.25% trypsin/EDTA (3 ml/dish, StemCell Technologies) and incubate for 3-5 min at 37 °C until the stem cells and PMEF detach from the plate. Add 15 ml fresh ES media without LIF and pipet cells up and down to break colonies (about 15-20 times). Then, add to a 0.1% gelatin-coated T150 flask and incubate for 30 min at 37 °C. After incubation, PMEF should attach to the gelatinized surface while ES cells should be floating. Collect the floating mES cells into a 50 ml tube.
- Spin down at 800 rpm and resuspend cells in 10 ml of Neural Differentiation Medium (see recipe in Table 1).
- Count the cells using a hemocytometer and then plate approximately 2 x 106 mES cells onto a 100-mm bacterial or suspension culture dish. These plates permit growth of suspension cultures and promote formation of embryoid bodies (EBs).
- On differentiation day 1, mES cells form small floating aggregates (EBs) that are visible under a light microscope. Any carryover PMEF attach to the dish. Transfer the suspension cells and media into a 15 ml tube and centrifuge at low speed (500 rpm) for 3 min. Aspirate media carefully, add 10 ml of fresh Neural Differentiation Medium to the pelleted EBs, and then plate cells in a new bacterial dish. In addition to replenishing the media, this step removes any PMEF that carry over from the prior step.
- On differentiation day 2, EBs are ready to be induced into motor neurons.
3. Step 3: Motor Neuron Specification (Timing: 5 days)
- On differentiation day 2, swirl the culture dish and transfer the media and EBs into a 15 ml tube. Let EBs settle by gravity (~ 10 min) or by centrifugation at low speed (500 rpm) for 3 min.
- Aspirate medium carefully and resuspend EBs in 10 ml of Motor Neuron Differentiation (MND) medium (see recipe in Table 1). Plate EBs in a new bacterial dish and change the media daily for 5 days.
- On differentiation day 7, EBs should be approximately 150 – 200 μm in diameter and should express strong GFP signals (Figure 1C). These EBs are ready to be dissociated and plated onto poly-DL-ornithine/laminin coated dishes or coverslips.
4. Step 4: Preparation of Poly-DL-ornithine/laminin Coated Coverslips (Timing: 2 days)
Two days before dissociating EBs (i.e. on differentiation day 5), prepare poly-DL-ornithine/laminin-coated coverslips.
- Place 12-mm round coverslips (Warner Instruments) on the bottom of a 24-well plate. Dissolve 25 mg poly-DL-ornithine (Sigma-Aldrich) in 25 ml sterile, double distilled water (d2H2O) to obtain a 10X stock solution (1 mg/ml). When ready to use, make a 1/10 dilution of poly-DL-ornithine with d2H2O to obtain concentration of 100 μg/ml. Add 0.5 ml to each well of the 24-well plate and incubate at 4 °C for overnight.
- The following day (differentiation day 6), aspirate poly-DL-ornithine and let the plates and cover slips air dry in a tissue culture hood. Rinse the plate wells with d2H2O three times. Let air dry for another hour.
- Dissolve 1 mg mouse laminin (Millipore) in 5 ml of iced-cold 1X PBS to make 100x stock solution (200 μg/ml). When ready to use, make 1/100 dilution in iced-cold 1X PBS (2 μg laminin/ml). Add 0.5 ml/well in a 24-well plate and incubate in 4 °C for overnight. Before seeding cells, excess laminin should be removed and wells rinsed with 1X PBS two times. Coverslips can be stored in MND medium (0.5 ml/well).
5. Step 5: Axon Elongation (Timing: 2 days)
- On differentiation day 7, collect EBs into a 15 ml tube and allow EBs to settle (approximately 3 min). Aspirate media and re-suspend EBs in PBS. Allow EBs to settle and then aspirate PBS. Repeat this step 3 times. Add 1 ml of Accumax (Millipore) to the EBs, gently mix and incubate for 5 min at room temperature. Then aspirate excess Accumax covering the EBs.
- To dissociate EBs, add 3 ml of MND medium. Pipet up and down 20 times using a 1 ml Eppendorf micropipette (blue tip) to disrupt EBs. Incubate for 2 min at room temperature. Transfer cell suspension to a 12×75 mm tube with cell strainer cap (BD Falcon) to obtain a single cell suspension. Repeat this dissociation step with remaining clumps and cells retained by the filter so as to enrich the yield of single cells. The final single cell suspension will be 6 ml.
- Gently mix this single cell suspension and count cells using a hemocytometer. Dilute cells into MND medium (supplemented with 10 ng/ml of each BDNF, GDNF, CNTF, and NT-3) to a density of 2×105 cells per ml. Add cell suspension (0.5 ml/well) to 24-well plates containing poly-DL-ornithine/laminin coated coverslips (previously prepared as described above). The differentiated cells extend long neurites after 2 days in culture (Figure 1D).
6. Representative Results
Figure 1A shows an outline of the protocol. In step 1, mES cells are cultivated on PMEF and supplemented with LIF to prevent spontaneous differentiation. In general, undifferentiated mES colonies are round and compact, and have clearly defined edges. The colonies are not in contact with one another. Typically, mES cells should be split at a 1:4 ratio every 2~3 days. However, each individual cell line will grow at a different rate and the split ratio must be determined empirically. Arrows in Figure 1B indicate the typical appearance of mES cells after culturing on a PMEF feeder layer for 3 days. These colonies are large (50-100 μm in diameter) but still maintain round and defined edges. At this stage the colonies are ready for differentiation or subculture. An arrowhead in Figure 1B shows a small mES colony that you typically find 24 h after plating. Four days after plating, mES cells become overgrown. They show a flattened appearance and loss of defined boundaries, indicating that they are beginning to differentiate. Such cells are not optimal for differentiation into neuronal cells.
To differentiate mES cells into motor neurons, mES cells need to be cultivated in suspension conditions to allow EB formation. In step 2, ES cells are transferred onto a culture surface without a feeder layer to promote EB formation. In this step, they are incubated in Neural Differentiation medium supplemented with Noggin, bFGF, and FGF-8 for 2 days to direct cells to a neural lineage. Small EBs can be observed under the microscope at day 1 of differentiation. They should be floating in the medium. Carryover PMEF can also be found at day 1 of differentiation and must be removed. These cells adhere to bacterial dishes so are eliminated when EBs are passaged to a new bacterial dish. Thus, by day 2 of differentiation, few or no PMEF should be seen in the culture dish. In step 3, continued transfer of EBs to new dishes should ensure removal of any remaining PMEF. EBs are cultured in Motor Neuron Differentiation (MND) media supplemented with RA and SAG for 5 days to differentiate the cells into motor neurons. During culture, EBs continue to grow in size and can be seen by the naked eye at differentiation day 3~4. Figure 1C shows the microscopic appearance of EBs on differentiation day 7. Unlike the undifferentiated cells, EBs show strong fluorescence due to expression of GFP. These EBs are optimally differentiated and are ready for dissociation. Flow cytometry showed that 51% ± 0.8% of the cells from the dissociated EBs had differentiated into GFP expressing cells5. In step 5, culture of these motor neurons in the presence of GDNF, CNTF, BDNF, and NT3 results in extension of long axonal projections. Figure 1D shows the appearance of differentiated cells 2 days after plating of dissociated EBs. Note long neurites extending from the cell bodies of the plated cells. Immunofluorescent staining shows that the GFP+ cells express the pan-neuronal marker (neurofilament-medium chain) and two motor neuron specific markers, (Islet-1 and choline acetyltransferase, Figure 2).
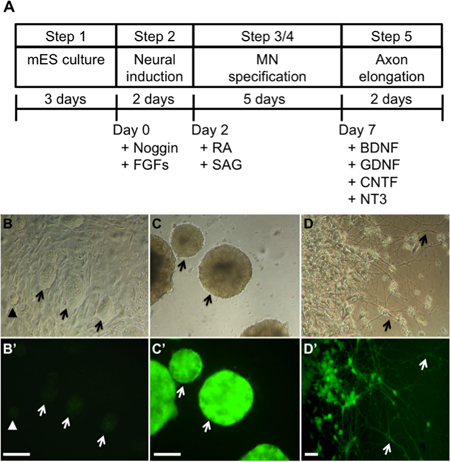
Figure 1. Differentiation of mES cells into motor neurons (picture modified from Wu et al.5). A) Scheme of the differentiation protocol from mES cells to motor neurons. B) Undifferentiated mES cells form round colonies on the top of a fibroblast feeder layer. They have weak GFP expression. C) mES cells were separated from feeder layers and cultured on low-attachment dishes to form EBs. During the first two days of the differentiation process, cells were exposed to 50 ng/ml Noggin, 20 ng/ml bFGF, and 20 ng/ml FGF-8. Subsequently, they were induced to differentiate with 1 μM retinoic acid and 1 μM sonic hedgehog agonist SAG for 5 days. Differentiated mES cells expressed strong GFP fluorescence. D) After 7-day differentiation, EBs were dissociated and plated on poly-DL-ornithine/laminin coated plates. The differentiated cells extended long neural processes after 2 days in culture. Scale bar = 200 μm. MN = motor neuron. B, C and D are bright field images and B’, C’ and D’ are corresponding fluorescence images.
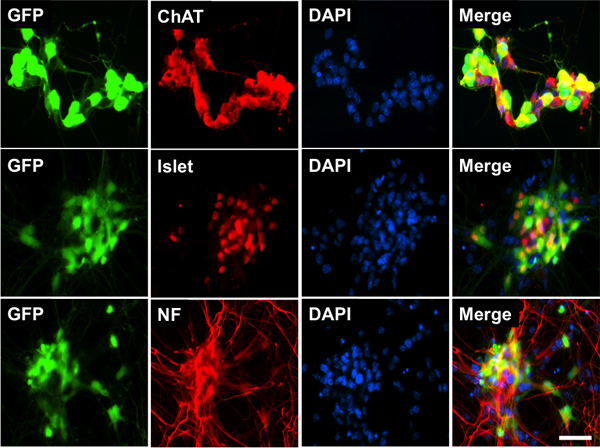
Figure 2. Characterization of mES cell-derived motor neurons. Immunofluorescence staining for neurofilament medium chain (NF-M, red), ChAT (red), and Islet-1 (red) was coincident with GFP (green) expression. DAPI (blue) was used to identify the nuclei. Scale bar = 50 μm.
