- Computer programs: Specialized computer programs are typically used in the generation of actograms and the calculation of circadian period. These programs include, but are not limited to, Actiview (Minimitter, Bend, OR) and Circadia.
- Actograms: Actograms provide a graphic illustration of the daily patterns of running-wheel activity. There are single-plotted (x-axis = 24 hr) and double-plotted (x-axis = 48 hr) actograms. Both methods plot sequential days from top to bottom, but double-plotted actograms plot two days on each horizontal line. Specifically, double-plotted actograms show the “second day” on the far right of each line, as well as at the start of the second horizontal line, and so on. Double-plotting is especially helpful to visualize non-24 hr rhythms.
- Periodogram: A periodogram is constructed from a spectral analysis of the running wheel activity over time. Periodograms show the relative power for a range of pre-set periods, and are commonly used to determine the free-running period.
- Results: In the laboratory, rodents are usually housed under a 24 hr LD cycle. Under these conditions the rhythm of activity is entrained, such that the daily pattern of running-wheel activity is aligned with the precise 24 hr LD cycle. In Figure 2A, the double-plotted actogram on the left shows the running-wheel activity of a rat that became active at the same time each day, soon after the environmental lights were turned off. The periodogram on the right shows a strong peak at 24 hr, consistent with entrainment to a precise 24 hr LD cycle. Figure 2B illustrates the running wheel use of a rat that was housed in constant darkness. In this case, the daily onset of running-wheel activity occurred slightly later each day, creating a rightward “drift”. This rightward “drift” indicates that the endogenous circadian period is greater than 24 hr, but it is the peak in the periodogram that quantifies this period. According to the periodogram, the maximum power is observed at 24.33 hr. In contrast to the automated periodogram analysis, Figure 3 illustrates a method to manually calculate the free-running period using the time of onset of running-wheel activity. It is important to realize that calculating the period by hand and calculating it with a periodogram may yield slightly different results.
The daily patterns of running-wheel activity can be disrupted in several ways. Figure 4 illustrates an arrhythmic pattern of running-wheel activity, produced by an electrolytic lesion of the SCN. This type of experiment provided some of the first evidence that the SCN contained the “master” circadian clock7,8. The periodogram on the right confirms this arrhythmic pattern of activity by showing equivalently low power for all periods in the circadian range (20-30 hr). The circadian pattern of running-wheel use can also be disrupted by housing rats in constant light. Figure 5 shows an actogram from a rat exposed sequentially to several of the lighting conditions already described. First, the rat was housed in constant darkness and exhibited a running wheel activity rhythm of approximately 24.33 hr. Second, the environmental light was kept on and the rat was housed in a constant light environment. Constant light is known to disrupt the SCN-based clock and produce arrhythmic patterns of running-wheel activity, similar to an SCN lesion. This disruption by light, however, occurs gradually over the course of 2-3 weeks. Therefore, when the running-wheel record is analyzed after the initial 3 weeks in constant light, the periodogram does not yield a peak. Finally, in the third phase the rat was put back on a 12 hr:12 hr LD cycle and the running wheel activity rhythms recover nearly immediately.
The amount of running, and the time of day it occurs, can also be manipulated by environmental factors. For instance, if rodents are fasted and given a temporally-restricted meal each day, this restricted feeding schedule will induce a daily bout of food-anticipatory activity. It is termed “anticipatory” because it occurs prior to the arrival of the daily meal, and is especially obvious when the meal is given in the middle of the day, a time when nocturnal rodents are relatively inactive. For instance, if an experiment provides a single 2 hr meal each day, food can be added to the cage at ZT 4 (4 hr after lights turned on) and removed at ZT 6 (2 hr later). Moreover, wire mesh flooring in the cages is also advantageous for this type of experiment because it makes it impossible for the rats to hide food and store it for later, thus ensuring that the rat is actually consuming all the food within the prescribed mealtime. Finally, one of the main advantages of an accurate running-wheel activity recording is that it allows for correlations to be made between running-wheel activity and daily oscillations in the expression of circadian clock gene expression throughout the brain and body. - Common pitfalls:
- Many software platforms automatically adjust for daylight saving time changes. When conducting an experiment at the time of yearly time changes, be sure this option is turned off in the recording software as well as the computer operating system software. This safeguard should help avoid discrepancies between the recording and the external light cycle.
- To be able to check on discrepancies in the data or unexpected changes in behavior, keep a text file with the precise time and date of every box opening, feeding and watering, bedding changes, experimental manipulations, and any other disruption that might occur. The precise start and end times of the data recording should also be noted in this file.
- It is important to check regularly that the lights are turning on and off at the expected times. Many problems can occur, including power outages and burnt out light bulbs. Some running-wheel platforms are equipped with light-sensors, but others do not verify lighting conditions.
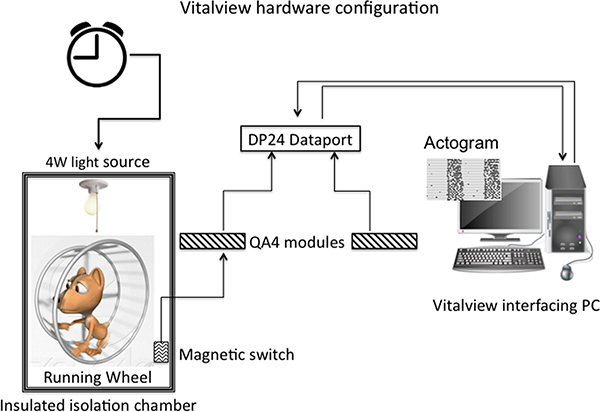
Figure 1. The Vitalview hardware configuration begins with a rodent running wheel, which is designed to activate a micro-switch with each revolution. This information then travels to the QA4 modules and is relayed to the DP24 dataport and finally is recorded by the Vitalview-equipped computer. The computer sums running-wheel revolutions from each channel every 10 min; these data can be viewed later as an actogram or periodogram. Depending on how the lights are set up, they can be controlled remotely either by the same Vitalview-equipped computer or by wall timers purchased from any electronics store.
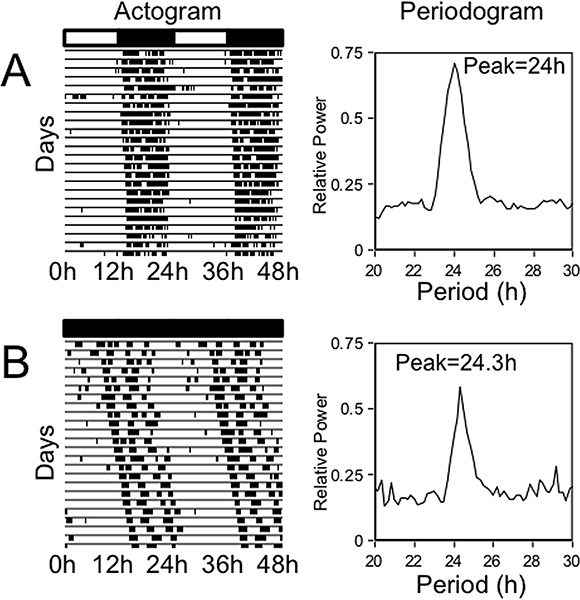
Figure 2. Representative actograms and periodograms for male Wistar rats housed in a 12 hr:12 hr LD cycle (A) and in constant darkness (B). Double-plotted actograms (left column) illustrate the lighting conditions along the top, 48 hr of running-wheel activity along the X-axis, and plot sequential days from top to bottom. Periodograms (right column) perform a spectral analysis on the running wheel data illustrated in the actograms. Figure 1A illustrates the behavior of a rat that was housed under a 24 hr LD cycle. Under these conditions the rat becomes active at the same time each day, showing an exact 24 hr peak in the periodogram. Figure 1B illustrates the behavior of a rat that was housed in constant darkness. Under these conditions the rat was active slightly later each day, hence the rightward drift in the actogram and 24.33 hr peak in the periodogram.

Figure 3. Period can also be extrapolated by hand. First draw a line of best-fit based on the daily onset of activity (red line). Next calculate the slope in h/day, remember that if the rhythm is <24 hr the slope will be a negative value, and finally add 24 hr. This procedure will provide an estimate of the circadian period for that animal. In this case, 4 hr/10 days suggests that the animal is becoming active approximately 0.4 hr later each day (slope 0.4 hr/day). Therefore, the circadian period is approximately 24.4 hr (or about 24 hr and 24 min). Any manipulation that is scheduled according to CT requires the free-running period to make an accurate prediction of CT.
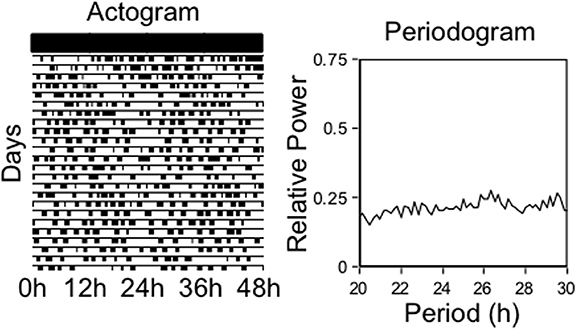
Figure 4. Electrolytic lesions of the SCN will produce arrhythmic patterns of running-wheel activity. In this case the rat is housed under constant darkness and, because the “master” circadian clock has been lesioned, the rat fails to show an endogenous circadian rhythm in running-wheel activity. The periodogram on the right confirms that there is no significant rhythm in the circadian range.
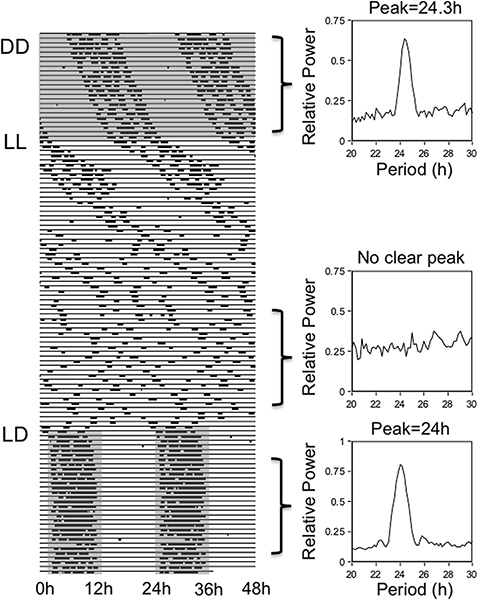
Figure 5. Lighting conditions have strong effects on the patterns of running wheel activity. In this record, the rat is initially housed under constant darkness (DD), as denoted by the shaded portion of the record. Under this condition, the circadian clock drives a circadian rhythm in running-wheel activity with a period of 24.33 hr, shown by the periodogram (top right). Next, the rat is housed under constant light (LL), as denoted by the white portion of the actogram. Under this condition the endogenous circadian clock is disrupted gradually over 2-3 weeks, and as shown by the periodogram (middle right) the rat becomes arrhythmic. Finally, the normal 12 hr:12 hr LD cycle was reinstated and the running-wheel activity rhythm was restored with a precise 24 hr rhythm as shown by the periodogram (bottom right).
