Integrations of transgenes in two different lines transmitting extrachromosomal arrays at about the same frequency (50-60%) were performed using the standard and the improved protocols (Table 1). The standard protocol allowed the recovery of three independent integrated lines after UV irradiation of 91 transgenic L4 animals, while the improved protocol allowed the recovery of one integrated line after UV irradiation of 30 transgenic L4 animals. This suggests that the efficiency of integration is similar for both protocols (one integrated line per 30 irradiated L4 animals). However, we cannot rule out the possibility that one protocol could be more efficient than the other when compared for the integration of the same transgene. The main difference between the protocols is that with the improved method all F1 plates exhibiting fluorescent progeny were selected regardless of the percentage of fluorescent F2 animals, while with the standard method a visual screen of F1 plates was performed and only plates with ≥75% transgenic F2 progeny were selected. This screening step is highly time consuming, in particular if the initial rate of transmission is close to 75%. In the improved protocol, it is sufficient to single four F2 worms from each F1 plate carrying transgenic F2 animals and to screen for plates with 100% transgenic progeny (F3), which can be performed quickly. In addition, the improved protocol allows for a decrease in the number of analyzed F2 and F3 animals when compared to the standard protocol. Altogether, the time and the number of plates necessary per integrated line are optimized in the improved protocol.
Next, we tested the hypothesis that a higher transgene transmission rate in a nonintegrated line renders easier the integration of the transgene into the genome. One particular transgene was integrated in lines transmitting at different rates using the improved protocol (Table 2). Gonads of wild type worms were injected with 10 ng/µl of a plasmid (pKG45) carrying the Pdyc-1::dyc-1::gfp transgene and with 30 ng/µl of the coinjection marker plasmid carrying Pmyo-2::dsRed10. Several transgenic strains were recovered and three of them that transmitted at different levels (10-15%, 50-60%, and >80%, respectively) were selected. The transgene was integrated in each strain using the improved protocol. The protocol was performed under the same condition for each strain, except that a higher number of transgenic L4 animals were irradiated for the line with the lowest transmitting rate (10-15%). For the highest transmitting line (>80%) three integrated lines were recovered from only 90 selected F1 animals, while for the transgenic line transmitting at 50-60%, one integrated line was recovered from 110 selected F1 animals. For the line with the lowest transmission rate, we could not recover a line carrying the integrated transgene, although 524 F1 animals were selected. Furthermore, the number of plates required per integrated line was three times lower with the highest transmitting line than with the line transmitting at 50-60% (Table 2).
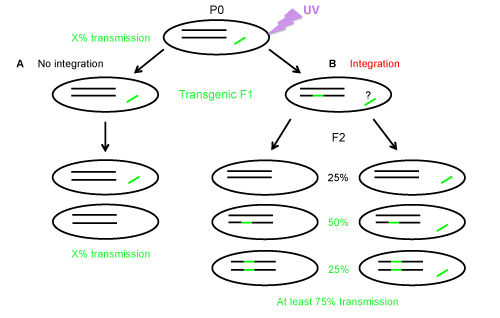
Figure 1. Transgene transmission in the progeny of transgenic worms with a nonintegrated or a heterozygous integrated transgene. A) Without integration, a transgenic animal will transmit the transgene at a frequency of X%, B) If integration of the transgene into a chromosome occurs; the F1 is most likely heterozygous for the integration and may carry or not the extrachromosomal array (as indicated by the question mark). The F2 progeny will be composed of at least 75% of fluorescent worms due to the Mendelian segregation. Worms without integrated transgene can be fluorescent, if they have retained the nonintegrated array (left column of Panel B).
Click here to view larger image.
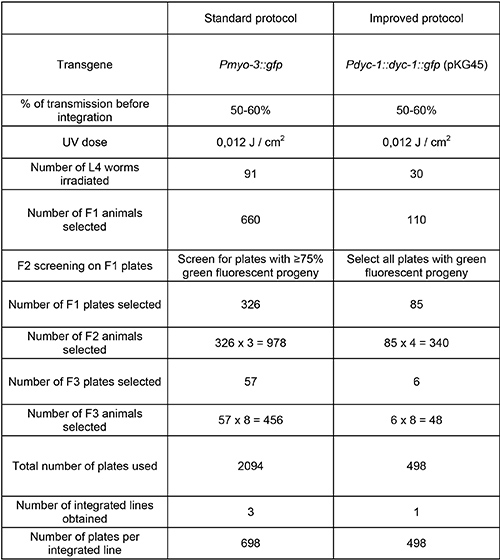
Table 1. Comparison of the standard and improved protocols for extrachromosomal arrays integration into the C. elegans genome. Two nonintegrated transgenes transmitting at about the same frequency were integrated using the standard or the improved protocol. The main difference lies in the fact that in the standard protocol a screen of the progeny on all F1 plates is performed to estimate the percentage of transgenic F2 worms, which should be ≥ 75%. This step takes a long time, particularly if the initial rate of transmission is close to 75%. In the improved protocol, it is sufficient to single four F2 worms from each F1 plate carrying transgenic F2 animals and to screen for 100% of transgenic worms in the progeny (F3), which can be performed quickly. The standard protocol is adapted from3.
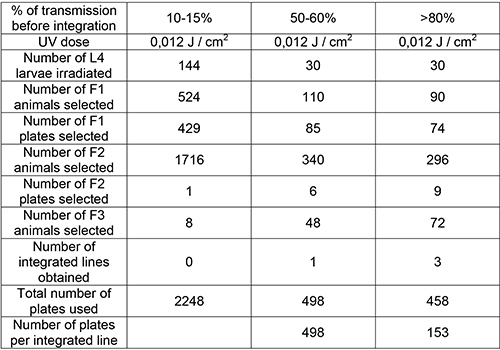
Table 2. Efficiency of transgene integration into the genome depends on the initial percentage of transgene transmission. Different nonintegrated lines, which transmitted the same transgene (Pdyc-1::dyc-1::gfp) at different rates (10-15%, 50-60%, and >80%, respectively) were used. Worms from each line were irradiated at the same UV dose (0.012 J/cm2) and the F3 generation screened for homozygous integration. The improved integration protocol applied to the highest transmitting line (>80%) recovered three integrated lines for only 74 selected F2 animals. The data presented with the line transmitting at 50-60% are those presented in Table 1 with the improved protocol.
