Culturing C. elegans in axenic liquid medium aids in the determination of nutrients that are required by worms, without interference from secondary metabolites produced by E. coli. Wildtype N2 worms acclimatize to mCeHR media within three generations and show growth comparable to worms grown on NGM bacterial plates. Indeed, these worms become gravid within 4 days as compared with 3.5 days for worms grown on OP50 bacteria.
One advantage of using mCeHR was seen in studies that examined the exact nutrient requirement of these worms14. In Figure 1 worms were grown in mCeHR supplemented with increasing amounts of heme, up to 1 mM. Observations of these worms showed a distinct delay in growth at heme concentrations below 4 μM, with worms developing to the L4 larval stage but unable to progress to the gravid stage after nine days in mCeHR. At concentrations of 10 μM and 20 μM heme the worms develop to the gravid stage in 4 days and produce large number of progeny. A maximum number of progeny was seen when worms were grown in mCeHR containing 20 μM heme. Worms continued to develop and produce progeny at 100 μM and 500 μM heme. However, the number of larval progeny significantly declined in comparison to worms grown at the optimal heme concentration of 20 μM heme. Heme concentrations at or above 800 μM resulted in stunted, sickly worms at the L3 larval stage, which indicated that these heme concentrations were toxic to the worms.
In addition to determining the optimal heme concentration and the effect of heme deprivation and heme toxicity on C. elegans growth, heme reporter strains could be utilized to indirectly assess the heme status of the worm within a smaller concentration range. The IQ6011 worm is a transgenic worm that expresses a heme responsive transcriptional reporter, Phrg-1::GFP, that inversely expresses GFP in response to environmental heme concentrations. When this worm is exposed to low environmental heme in mCeHR, GFP is highly expressed. This response is reversed under heme replete conditions, as seen in Figure 2. GFP expression is repressed at 20 μM heme and increases as the heme concentration is decreased. The incremental changes in heme concentration can be accurately correlated with gene response and expression in mCeHR media.
In addition to being able to carefully control nutrient concentrations provided to the worm, mCeHR axenic media allows for the efficient growth of a large number of synchronized worms. This feature can be exploited for microparticle bombardment (Figure 3). Using this procedure at least one integrated line have been developed from every microparticle bombardment carried out (Table 2).
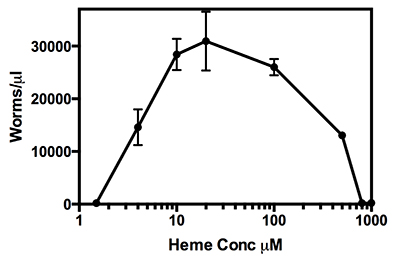
Figure 1. C. elegans heme growth curve. Worms were grown in a range of heme concentrations from 0 μM to 1,000 μM in mCeHR for 9 days. The number of worms in each concentration was counted and plotted. At 1.5 μM heme concentrations worms were L4 and unable to further develop. At 800 μM heme the worms were stunted at L2-L3 stages and showed effects of heme toxicity.

Figure 2. Response of heme sensor (Phrg-1::GFP) strain to different heme concentrations in mCeHR. Synchronized transgenic C. elegans expressing hrg-1::GFP were grown in mCeHR media supplemented with 4, 8, 10, or 20 µM heme for 48 hr. Images were taken with a Zeiss LSM 710 confocal microscope. Scale bar is 100 µm. Please click here to view a larger version of this figure.
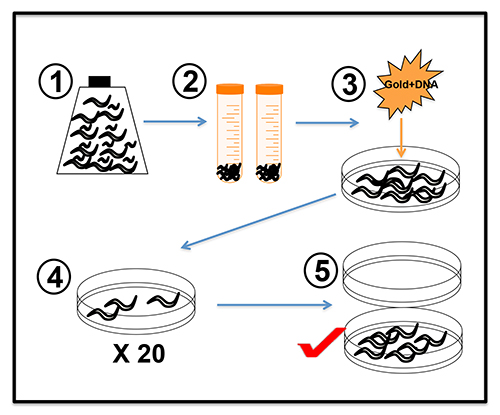
Figure 3. Schematic of microparticle bombardment in C. elegans using unc-119 worms grown in mCeHR. (1) Approximately 3 x 107 unc-119(ed3) worms are grown in 90 ml mCeHR media. (2) Gravids were allowed to settle on ice for 10-15 min in 50 ml conical tubes. (3) A 2 ml pellet of approximately 5 x 106 gravid worms was spread evenly onto an unseeded NGM plate. The worms were bombarded with 12 μg of plasmid of interest and 6 μg of unc-119 rescue plasmid complexed to gold particles. (4) The bombarded worms were split onto twenty 10 cm plates seeded with the E. coli strain JM109. (5) After 2 weeks incubation at 25 °C, plates with wild type worms were selected for analysis of transgene expression strength and segregation rates.
| Table 1A | |
| CeHR, 1 L | |
| Using sterile technique and a 1 L (0.22 μm) vacuum filter unit, filter the following volumes of stock solutions and water in the order described. | |
| Choline diacid citrate | 10 ml |
| Vitamin and growth factor mix | 10 ml |
| myo-Inositol | 10 ml |
| Hemin chloride | 10 ml |
| Deionized water | 250 ml |
| Nucleic acid mix | 20 ml |
| Mineral Mix | 100 ml |
| Lactalbumin hydrolysate | 20 ml |
| Essential Amino Acid Mix | 20 ml |
| Non-essential Amino Acid Mix | 10 ml |
| KH2PO4 | 20 ml |
| D-Glucose | 50 ml |
| HEPES, sodium salt | 10 ml |
| Deionized water | 250 ml |
| Volume will be 800 ml at this point Remove filter unit from vacuum then add: |
|
| Cholesterol | 1 ml |
| Ultra-pasteurized skim milk | 200 ml |
| Table 1B | |
| Vitamin and growth factor mix, 100 ml | |
| Solution 1: To 60 ml of water add: | |
| N-acetyl-α-D-glucosamine | 0.15 g |
| DL-alanine | 0.15 g |
| Nicotinamide | 0.075 g |
| D-pantethine | 0.0375 g |
| DL-pantothenic acid, hemi calcium salt | 0.075 g |
| Folic acid | 0.075 g |
| Pyridoxamine 2HCl | 0.0375 g |
| Pyridoxine HCl | 0.075 g |
| Flavin mononucleotide, sodium salt | 0.075 g |
| Thiamine hydrochloride | 0.075 g |
| Solution 2: Prepare the following chemicals in 5 ml 1 N KOH: | |
| p-aminobenzoic acid | 0.075 g |
| D-biotin | 0.0375 g |
| Cyanocobalamin (B12) | 0.0375 g |
| Folinic acid, calcium salt | 0.0375 g |
| Nicotinic acid | 0.075 g |
| Pyridoxal 5-phosphate | 0.0375 g |
| Solution 3: 0.0375 g (±) α-L-lipoic acid, oxidized form in 1 ml ethanol: | |
| Combine solutions 1, 2, and 3 and bring the final volume to 100 ml. Store in dark at 4 °C or freeze aliquots at -20 °C. Make small volumes of stocks for this mix so it is used quickly. |
|
| Table 1C | |
| Nucleic acid mix, 100 ml | |
| To 60 ml of water add: | |
| Adenosine 5' -monophosphate, sodium salt | 1.74 g |
| Cytidine 5' -phosphate | 1.84 g |
| Guanosine 2' – and 3' -monophosphate | 1.82 g |
| OR | |
| Guanosine 5' -phosphate | 2.04 g |
| Uridine 5' -phosphate, disodium salt | 1.84 g |
| Thymine (add last) | 0.63 g |
| Bring solution to 100 ml and store in the dark at 4 °C or freeze aliquots at -20 °C. Make small volumes of stocks for this mix so it is used quickly. |
|
| Table 1D | |
| Mineral Mix, 1 L | |
| MgCl2•6H2O | 4.1 g |
| Sodium citrate | 2.9 g |
| Potassium citrate monohydrate | 4.9 g |
| CuCl2•2H2O | 0.07 g |
| MnCl2•4H2O | 0.2 g |
| ZnCl2 | 0.1 g |
| Fe(NH4)2(SO4)2•6H2O | 0.6 g |
| CaCl2•2H2O (always add last) | 0.2 g |
| Make small volumes of stocks for this mix so it is used quickly. | |
| Table 1E | |
| Other Components | |
| KH2PO4 | 450 mM |
| Choline di-acid citrate | 2 mM |
| myo-Inositol | 2.4 mM |
| D-Glucose | 1.45 M |
| Hemin chloride | 2 mM in 0.1 N NaOH pH 8.0 |
| HEPES, sodium salt | 1 M stock solution |
| Cholesterol | 5 mg/ml in ethanol |
| Lactalbumin enzymatic hydrolysate | 170 mg/ml |
| Table 1F | |
| M9 Buffer, 1 L | |
| KH2PO4 | 3 g |
| Na2HPO4 | 6 g |
| NaCl | 5 g |
| H2O | 1 L |
| Autoclave 30 min | |
| 1 M MgSO4 (sterile) | 1 ml |
Table 1. Recipes for components of mCeHR and mCeHR.
| Bombardment | Lines with wild type rescue | Lines with rescue / transgene | Stable lines |
| 1 | 2 | 2 | 1 |
| 2 | 8 | 5 | 0 |
| 3 | 4 | 4 | 2 |
| 4 | 5 | 2 | 1 |
| 5 | 5 | 3 | 1 |
| Average | 4.8 | 3.2 | 1 |
Table 2. Average number of transgenic C. elegans generated using microparticle bombardment.
