A partir do "mosaico fluido" modelo original por Singer e Nicolson, a imagem da membrana plasmática celular tem sido continuamente atualizado durante as últimas décadas, a fim de incluir o papel emergente do citoesqueleto e lipídios domínios 1,2.
As primeiras observações foram obtidos por recuperação de fluorescência após a fotodegradação (FRAP) revelação de que uma fração significativa de proteínas de membrana é imóvel 3-5. Estes estudos pioneiros, embora muito informativo, sofreu com a relativamente baixa resolução no espaço (microns) e de tempo (segundos) de setups FRAP. Além disso, sendo uma medição de média conjunto, FRAP falta em dar informações sobre o comportamento única molécula.
Neste contexto, a possibilidade de marcar especificamente uma única molécula com marcações muito brilhantes (permitindo o estudo do processo de difusão de uma molécula de cada vez), tem sido muito bem sucedida. Particularmente, empurrando otempo de resolução da abordagem única partícula Tracking (SPT) para a escala de tempo microssegundos, Kusumi, et al. acesso ganhou a recursos desconhecidos de lipídeos e proteínas dinâmica que muito contribuíram para o reconhecimento do papel do esqueleto da membrana à base de actina na fisiologia da membrana 6 , 7. Essas descobertas geraram a chamada do "piquete e cerca 'modelo, em que lipídios e proteínas difusão é regulada por esqueleto à base de actina. No entanto, a fim de ter acesso à enorme quantidade de informações fornecidas pelo SPT muitas questões experimentais têm de ser abordadas. Particularmente, o processo de marcação é tipicamente composta por várias etapas, como produção, purificação e introdução das espécies marcadas no sistema. Além disso, grandes gravadoras, como pontos quânticos ou nanopartículas de metal, muitas vezes são necessárias para alcançar a escala de tempo sub-milissegundo ea reticulação das moléculas alvo pelo rótulo não pode ser evitado em muitos casos. Finalmente, muitas trajetóriaster de ser de forma a caber critérios estatísticos e concomitantemente uma baixa densidade da etiqueta é necessária para permitir o rastreamento.
Comparado a SPT, espectroscopia de correlação de fluorescência (FCS), superando muitos destes inconvenientes, representa uma abordagem muito promissora para o estudo de dinâmica molecular. Na verdade, FCS funciona bem também com rótulos escuros e densos, permitindo estudar a dinâmica das moléculas marcadas em proteínas fluorescentes em células transfectadas transitoriamente. Além disso, permite alcançar altas estatísticas em uma quantidade limitada de tempo. Por fim, apesar da "alta" densidade de rótulos FCS fornece moléculas únicas informações. Graças a todas essas propriedades, FCS representa uma abordagem muito simples e tem sido amplamente utilizada no estudo da dinâmica de lipídeos e proteínas, tanto em membranas modelo e em células vivas 8-10. Muitas abordagens diferentes têm sido propostos para aumentar a capacidade de FCS para revelar os detalhes de difusão molecular. Por exemplo, foi shprópria, que através da realização FCS em áreas de observação de diferentes tamanhos pode-se definir uma "lei de difusão FCS" recursos escondidos esclarecedoras do movimento molecular 11,12. Além de ser variaram em tamanho, a área focal também foi duplicada 13, mudou-se no espaço ao longo das linhas 14-20 ou conjugadas com câmeras rápidas 21,22. Usando estes correlação 'espaço-temporal' abordagens, os parâmetros biológicos relevantes para vários componentes da membrana foram quantitativamente descrito em ambas as membranas modelo e as biológicas reais, portanto, uma visão rendimento em membrana de organização espacial.
No entanto, em todas as aplicações e FRAP FCS descrito até agora, a dimensão da área focal representa um limite de resolução espacial, em que não podem ser ultrapassados. Vários métodos de imagem de super-resolução foram recentemente desenvolvidos para contornar este limite. Alguns são baseados em precisão de localização, tais como microscopia óptica reconstrução estocástica (STORM) <sup> 23,24, microscopia fotoativação localização (PALM) 25, PALM fluorescência (FPALM) 26 e de uma única partícula de rastreamento PALM (sptPALM) 27: a quantidade relativamente grande de fótons necessários em cada snapshot, no entanto, limita o tempo de resolução de estes métodos para, pelo menos, vários milissegundos, prejudicando assim a sua aplicabilidade in vivo.
Em contraste, uma alternativa promissora para a resolução de imagem de super ter sido aberto por espacialmente modulando a emissão de fluorescência com os métodos de depleção de emissão estimulada (STED ou reversíveis transições saturáveis óptica de fluorescência (RESOLFT)) 28,29. Estas abordagens combinar a formação de um volume de observação bem abaixo do limite de difracção, com a possibilidade de utilização de microscópios varrimento rápido e sistemas de detecção. Em combinação com a análise de variação de fluorescência, microscopia STED permissão para investigar diretamente a dinâmica espaço-temporais em nanoescala de lipídeos e proteins em membranas de células vivas 30,31.
As mesmas quantidades físicas de microscopia-STED base pode ser obtida por uma espectroscopia modificado espaço-temporal correlação imagem (STICS 32,33) método que é adequado para o estudo da dinâmica das proteínas marcadas com fluorescência de membrana e / ou lípidos em células vivas e por um microscópio comercial. O protocolo experimental aqui apresentada é composta por poucos passos. O primeiro exige uma imagem rápida da região de interesse na membrana. Em seguida, a pilha resultante de imagens é usada para calcular as funções de correlação espaço-temporais médios. Ajustando-se a série de funções de correlação, a molecular "lei difusão" pode ser obtida directamente a partir de imagens na forma de uma difusividade aparente (D app) – vs -Média plotagem de deslocamento. Este lote depende criticamente sobre o meio ambiente explorado pelas moléculas e permite reconhecer diretamente os modos de difusão reaisdo lípido / proteína de interesse.
Em mais detalhes, como demonstrado anteriormente 34, a função de auto-correlação espaço-temporal da série imagem adquirida de forma crítica depende da dinâmica das moléculas em movimento na série de imagens coletadas (por favor, note que o mesmo raciocínio pode ser aplicado em uma aquisição de linha onde apenas uma dimensão do espaço é considerado). Em particular, podemos definir a função de correlação como:
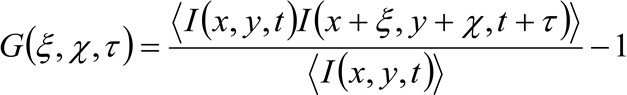 (1)
(1)
onde 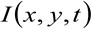 representa a intensidade da fluorescência medida, na posição x, y e no momento t,
representa a intensidade da fluorescência medida, na posição x, y e no momento t, ![]() e
e ![]() representa a distância em x edireções y respectivamente,
representa a distância em x edireções y respectivamente, ![]() representa o intervalo de tempo, e
representa o intervalo de tempo, e ![]() representa a média. Esta função pode ser expressa como:
representa a média. Esta função pode ser expressa como:
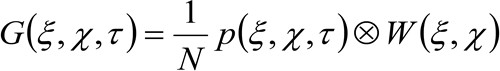 (2)
(2)
onde "N" representa o número médio de moléculas na área de observação, ![]() representa a operação de convolução no espaço, e
representa a operação de convolução no espaço, e 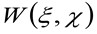 representa a autocorrelação da cintura instrumental. Este último pode ser interpretada como uma medida de como os fotões de um único emissor estão espalhados no espaço, devido à configuração óptica / gravação (o chamado ponto de propagação função, PSF, generali bem aproximada por uma função de Gauss). Finalmente,
representa a autocorrelação da cintura instrumental. Este último pode ser interpretada como uma medida de como os fotões de um único emissor estão espalhados no espaço, devido à configuração óptica / gravação (o chamado ponto de propagação função, PSF, generali bem aproximada por uma função de Gauss). Finalmente, 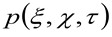 representa a probabilidade de encontrar uma partícula a uma distância
representa a probabilidade de encontrar uma partícula a uma distância ![]() e
e ![]() depois de um tempo de atraso
depois de um tempo de atraso ![]() . Se considerarmos uma dinâmica difusos, em que as partículas se movem aleatoriamente em todas as direções e fluxos líquidos não estão presentes, esta função também é bem aproximada por uma função de Gauss, onde a variância pode ser identificado como o Mean Square Deslocamento (MSD) da partícula em movimento . Assim, a cintura da função de correlação (também referida como
. Se considerarmos uma dinâmica difusos, em que as partículas se movem aleatoriamente em todas as direções e fluxos líquidos não estão presentes, esta função também é bem aproximada por uma função de Gauss, onde a variância pode ser identificado como o Mean Square Deslocamento (MSD) da partícula em movimento . Assim, a cintura da função de correlação (também referida como ![]() ), Pode ser definido como a soma das perturbações músculo-esqueléticas e de partículas da cintura instrumental e pode ser medido por um ajuste de Gaussção da função de correlação, para cada intervalo de tempo. A medida MSD i pode ser utilizada para calcular uma difusividade aparente das moléculas que se deslocam
), Pode ser definido como a soma das perturbações músculo-esqueléticas e de partículas da cintura instrumental e pode ser medido por um ajuste de Gaussção da função de correlação, para cada intervalo de tempo. A medida MSD i pode ser utilizada para calcular uma difusividade aparente das moléculas que se deslocam ![]() e uma média de deslocamento
e uma média de deslocamento ![]() como:
como:
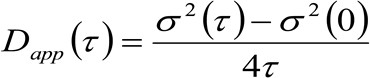 (3)
(3)
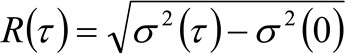 (4)
(4)
Algumas considerações sobre a configuração experimental utilizada pode guiar o leitor ao longo das próximas seções. A fim de estimular selectivamente os fluoróforos na membrana basal de células que irão utilizar uma iluminação de reflexão interna total (TIR), vivo, usando uma fluorescência TIR comercial (TIRF) microscópio (detalhes podem ser encontrados na secção de material). Além disso, a fim de recolher the fluorescência usaremos um objectivo ampliação elevada (100X NA 1,47, abertura numérica elevada é necessária para a iluminação TIRF) e uma câmara EMCCD (tamanho físico do pixel no chip 16 mm). Para chegar a um tamanho de pixel de 100 nm, aplicamos uma lente de ampliação adicional de 1.6X. Como discutido abaixo, uma resolução temporal inferior a 1 ms seria necessário para descrever adequadamente a dinâmica dos lípidos da membrana rápidos abaixo de 100 nm. Para chegar a esta resolução temporal, precisamos selecionar uma região de interesse (ROI) menor do que todo o chip da câmera (512 x 512). Desta forma, a câmara irá ler um número reduzido de linhas de aumentar a resolução de tempo. No entanto, neste regime de tempo de leitura do quadro seria limitada pelo tempo necessário para mudar as taxas de exposição ao chip de leitura na câmara e é geralmente da ordem de milissegundos para 512 x 512 pixels EMCCD. Para vencer esse limite, uma tecnologia emergente permite deslocar as linhas de ROI só que ao invés de todo o quadro, wom uma redução efetiva prática do tamanho do chip exposto (chamado Modo Sensor recortada em nosso EMCCD). Para esta configuração para ser eficaz, o chip de fora do ROI deve ser coberta por um par de fendas montadas no caminho óptico. Graças a esta configuração de uma resolução de tempo até 10 segundos -4 pode ser alcançado. Observe, porém, que esta abordagem pode ser acoplado com muitas montagens experimentais diferentes, como explicado na seção "discussão".
Demonstração do método irá ser fornecida em células vivas, utilizando tanto uma ATTO488 identificada como 1-palmitoil-2-hidroxi-sn-glicero-3-fosfoetanolamina (ATTO488-PPE) e uma variante marcada com GFP do Receptor de Transferrina (GFP TFT). No caso de ATTO488-PPE esta abordagem pode recuperar com sucesso um aplicativo D quase constante como uma função da média de deslocamento que indica uma difusão maior parte livre, como relatado anteriormente 30,35. Por contraste, o TfR-GFP demonstra uma diminuição D <sub> app em função do deslocamento da média, sugerindo difusão parcialmente confinado 6. Além disso, neste último caso, é possível quantificar a constante de difusão local, e a média da área ao longo de muitos micra confinamento no plano da membrana.
