In Figure 1 is visualized the investigation and quantification of hNSC differentiation on 3D scaffolds compared with traditional flat surface cultures (2D) and expressedas axon process outgrowth measurements. In Figure 2 is representedthe workflow and the results for the miRNA quantification using stem cells and developmental pathways focused miRNA PCR array to investigate differential expression of miRNAs in 2D and 3D differentiated hNSCs compared with undifferentiated control. Following miRNA comparative analysis between differentiated and undifferentiated hNSCs, and computational analysis for target prediction, SOX5 and NR4A3 (NOR1) were identified as putative target mRNAs. To study the direct interaction between the miRNAs and their putative site on the 3’UTR mRNA we used 2 commercially available plasmids containing either SOX5 or NR4A3 3′UTR sequence inserted downstream of the firefly luciferase reporter gene, and containing in the same plasmid renilla luciferase gene for normalization. The representative results of 3’ UTR luciferase activity down-regulation is presented in Figure 3.
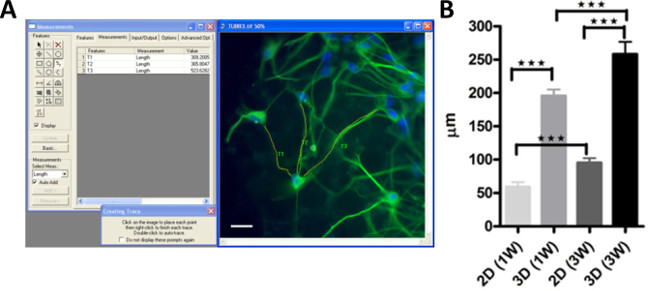
Figure 1: Differentiation of hNSCs on 3D scaffolds, and measurement of axon processes outgrowth. (A) Following standard ICC staining for β3-tubulin (green) and nuclei counterstained with Hoechst (blue) representative microphotographs were acquired and measurement of axon processes were performed with the aid of an image analysis software. (B) Diagrams reporting the measurement of axon process outgrowth performed in 2D and 3D differentiated hNSCs for 1 (1W) or 3 weeks (3W). Please click here to view a larger version of this figure.
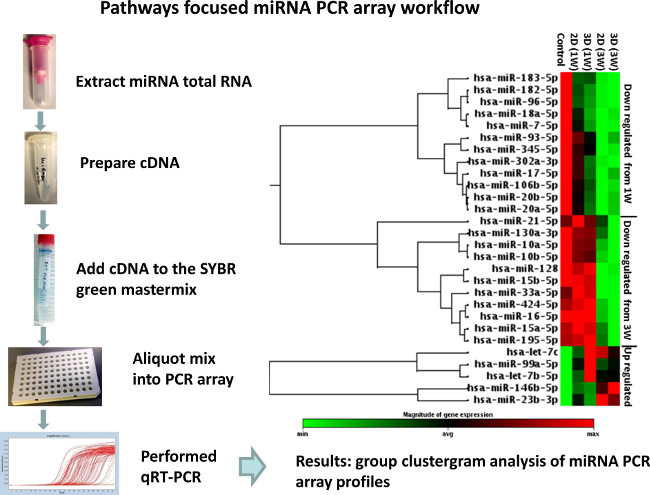
Figure 2: Schematic workflow of miRNA profiling in differentiated hNSCs. Total RNA,including miRNAs, was extracted from either differentiated hNSCs on 3D scaffolds and standard 2D cultures, or undifferentiated control. 250 ng of total RNA were retro-transcribed with a method that facilitates the selective conversion of mature miRNAs into cDNA suitable for miRNA quantification with human cell differentiation and development miScript miRNA PCR array. The geo-mean of SNORD61, SNORD68, SNORD72, SNORD95, SNORD96A, and RNU6-2 was used for data analysis based on the 2-ΔΔct method. Significant changes were defined as +/- 1.5 fold up and down regulation to a statistically significant extent. Data were analyzed using web-based software available at Results are expressed as a clustergram. Modified from Figure 319. Please click here to view a larger version of this figure.
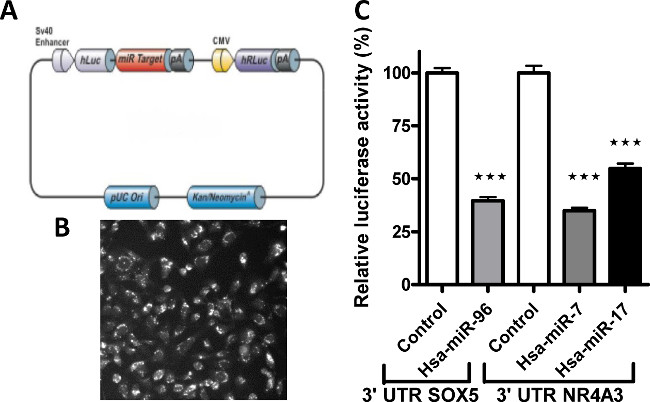
Figure 3: Validation of miRNA target predicted mRNA using a dual luciferase report assay. (A) Representative diagram of a dual luciferase reporter plasmid used as a tool to validate 3’UTR sequence as a direct target of a miRNA. (B) Representative micrograph showing transfection efficiency. (C) Signals, expressed as relative luciferase activity, from human 3'UTR reporters ofSOX5 and NR4A3 genes were significantly knocked down in the presence of hsa-miR-96, and hsa-miR-7 and 17 miRNA mimics respectively. Modified from Figure 519. Please click here to view a larger version of this figure.
| DIANALAB | PicTar | TargetScan | |||
| miRNA | Target gene | Target sites/genes found | Precision | Score | Aggregate PCT |
| hsa-miR-96 | SOX5 | 1214/763 | 1 | 6.74 | 0.94 |
| hsa-miR-183 | DUSP10 | 302/190 | 0.72 | 4.75 | 0.85 |
| hsa-miR-302a | NR4A3 | 863/473 | 0.84 | 3.05 | NF |
| hsa-miR-182 | FOXN3 | 1358/794 | 0.94 | NF | 0.96 |
| hsa-miR-7 | NR4A3 | NF | NF | 1.61 | 0.17 |
| hsa-miR-7 | FOXN3 | 399/136 | 0.17 | NF | 0.88 |
| hsa-miR-20a | NR4A3 | 1650/841 | 0.85 | 5.87 | 0.63 |
| hsa-miR-20b | NR4A3 | 1955/973 | 0.9 | 5.87 | 0.63 |
| hsa-miR-17 | NR4A3 | 1928/961 | 0.9 | 5.38 | 0.63+0.37 |
| hsa-miR-20a | EIF4G3 | 1650/841 | 0.97 | NF | NF |
| hsa-miR-20b | EIF4G3 | 1955/973 | 0.94 | NF | NF |
| hsa-miR-17 | EIF4G3 | 1928/961 | 0.94 | NF | NF |
Table 1: List of miRNA target prediction mRNAs. Representative table of miRNAs, target predictive genes,and algorithms analysis (prediction/score/aggregate provided using Diana Lab, PicTar, and TargetScan respectively). Modified from Table 319.
