T الليمفاوية هي فرع من نظام المناعة التكيفية التي تتميز القدرة على التعرف على كفاءة الببتيد المستضد (حج) منضمة إلى رئيسي معقد التوافق النسيجي (MHC) الجزيئات من خلال مستقبلات الخلايا T الخاصة بهم (TCRs) 1. اللمفاويات السذاجة تهاجر بشكل جوهري ومسح "حج تقديم الخلايا المهنية" (ناقلات الجنود المدرعة، على سبيل المثال، والخلايا الجذعية) داخل الغدد الليمفاوية، بينما تحتاج خلايا الذاكرة / T المستجيب لدراسة فعالية مجموعة واسعة جدا من ناقلات الجنود المدرعة والخلايا المستهدفة المحتملة داخل الأنسجة الطرفية.
في دقيقة بعد الاعتراف الأولي وما شابه ذلك حج على APC، الخلايا اللمفية اعتقال هجرتهم والبدء في تشكيل الحميمة واجهة خلية خلية متخصصة تسمى "المشبك المناعي" (IS). أصيب (أي 30-60 دقيقة) هو يطلب من الاتصالات لتضخيم والحفاظ يشير 2-7. دراسات الناشئة تحديد ذلك ضمن IS، هو تشكيل المستمر وص السريعemodeling من الإشارات المنفصلة شبه الخلوية مجموعات صغيرة (أي التي تحتوي على MHC / حج-TCR، F-الأكتين، التصاق والجزيئات يشير) التي تحدد قوة وجودة الناتج الاستجابات المناعية 2-7. ومع ذلك، تفاصيل ديناميكية وآلية تنظيمية لهذه العملية هي غير مفهومة تماما 8،9. هذا نابع إلى حد كبير من التحديات الفنية المرتبطة طبولوجيا عدم انتظام السطوح APC والتوجه سيئة تسيطر الطائرات التفاعل خلية خلية، المسائل التي تحد عميقا في التصوير الزمانية المكانية اللازمة النهج 10/08 (Figure1A).
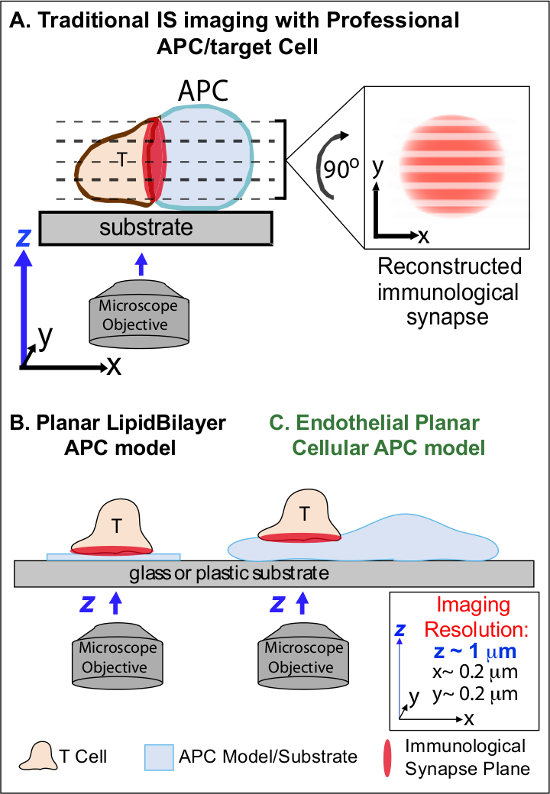
الشكل 1. فسيولوجي مستو نموذج APC خلية التصوير المناعية سنبس حيوية. ويوضح التخطيطي التصوير التقليدي للالمشبك المناعي بين الخلايا التائية والمهنيه نال APC (A) والخلايا التائية ومستو الدهون النموذج التقليدي طبقة ثنائية APC (B) بالمقارنة مع هذه البطانية نموذج مستو APC رواية (C). توفر ناقلات الجنود المدرعة المهنية نقاط الاشتباك العصبي المناعية الفسيولوجية ولكن تقدم سيئة موجهة اجهة خلية خلية (أي فيما يتعلق طائرة التصوير س ص المثلى؛ والقرار ~ 0.2 ميكرون)، والذي يعرض للخطر بشكل كبير المكاني (طائرة التصوير ض قرار ~ 1 ميكرون)، والزمانية (أي، بسبب الحاجة لمسح مرارا وتكرارا من خلال جميع الطائرات ض التصوير) قرار من التصوير. نماذج طبقة ثنائية لديها طوبولوجيا مستو الذي يوفر الأمثل القرار التصوير الزمانية المكانية، ولكن أيضا مبسطة للغاية، غير الفسيولوجية وجامدة. هذا النموذج الخلايا البطانية يجمع بين طوبولوجيا مستو من طبقات ثنائية الدهون مع الركيزة الفسيولوجية من APC الكلاسيكية لتقديم الأمثل القرار التصوير المكاني والزماني في وضع فيزيولوجي.م / ملفات / ftp_upload / 53288 / 53288fig1large.jpg "الهدف =" _ فارغة "> الرجاء انقر هنا لعرض نسخة أكبر من هذا الرقم.
العمل السابق والتحايل جزئيا على هذه العقبات من خلال تطوير نماذج مستو الركيزة (أي طبقات ثنائية الدهون والأسطح المغلفة الأجسام المضادة) التي توفر قرار الزمانية المكانية الأمثل (أي عن طريق تحديد T تنشيط الخلايا السطحية في خطة واحدة التي هي موازية لتصوير س ص الأمثل الطائرة) 11-15 (الشكل 1B). وقد سهلت هذه النماذج معلومات هامة في ديناميات التحت خلوية / الجزيئية التي تتحكم في الإشارات المستضدات في الخلايا T، بما في ذلك اكتشاف الأكتين ديناميكية / TCR يشير-مجموعات الصغيرة 7،11-14. ومع ذلك، فإن التبسيط بطبيعتها مثل هذه النماذج، وكذلك جامدة (النافية للتنمية / دراسة الخصائص الطبوغرافية 3-الأبعاد) (الشكل 1B). ولذلك، فإنه لا يزال غير مؤكد كيفية ربط هذه النتائج إلى فيزsiologic خلية خلية المراقبة المناعية.
وإن كان لا يزال دراسة سلوكه، الأوعية الدموية والخلايا البطانية اللمفاوية آخذة في الظهور كما كبيرة (أي أكبر في أعداد من جميع ناقلات الجنود المدرعة المهنية، التي كتبها ~ 1000 مرات) حجرة الطرفية من "شبه المحترفين ناقلات الجنود المدرعة 16-18. هذه الخلايا تعبر عن MHC-I-، MHC-II- وعدد كبير من الجزيئات المشارك مشجعا (على سبيل المثال، CD40، LFA3، ICOSL، 4-1BB، OX40L، TL1A، PD-L1، ولكن لا CD80 CD86 و) وهي استراتيجيا وضعه في واجهة الأنسجة الدموية التي يعملون فيها وظائف الحارس المتخصصة 16-18. وأظهرت دراسات سابقة أن الخلايا البطانية على نحو فعال إعادة تحفيز المستجيب / الذاكرة، ولكن ليس ساذجا، وخلايا T 19-25. وبالتالي، من المرجح أن يلعب أدوار APC فريدة من نوعها في المرحلة المستجيب من الاستجابات المناعية التكيفية داخل الأنسجة الطرفية، مثل التأثير المحلي على T تنشيط الخلايا، والتمايز، والذاكرة والتسامح 16،17،26 الخلايا البطانية. CRItically، عندما نمت في المختبر، الخلايا البطانية شكل سطوح الخلايا مستو تقريبا وهي transfectable بسهولة (على سبيل المثال، مع الصحفيين بروتين فلوري). هذه الميزات هي مثالية عالية الدقة التصوير الزمانية المكانية لديناميات الطوبوغرافية خلال التفاعلات خلية خلية 19،27. وهكذا الخلايا البطانية قد تكون بمثابة فيزيولوجي "مستو الخلوي APC" النموذجية مناسبة مميزة لدراسة آليات إعادة التحت خلوية / الجزيئية التي تدفع الاعتراف مستضد وتنظيم الاستجابات (الشكل 1C) 19،20.
أنشئت سابقا تقنيات التصوير التكميلية (بما في ذلك ترنسفكأيشن من خلايا بطائن مع صانعي بروتين فلوري من غشاء البلازما والعصارة الخلوية) لدراسة تفاصيل تفاعل الكريات البيض البطانية خلال الالتصاق والهجرة transendothelial 27، أظهرت أن الكريات البيض تسبر بنشاط سطح البطانة التي كتبها ديناميكية الإدراج فيد تراجع من ميكرون نطاق الفرعي، نتوءات أسطواني الأكتين الغنية (~ 200-1،000 نانومتر في القطر والعمق) يطلق مثل invadosome نتوءات (أي، "ILPs ') 27،28. تم زيادة توسيع هذه النهج التصوير جنبا إلى جنب مع إنشاء بروتوكولات للاستفادة من البطانية وظيفة APC لتطوير أساليب الأولى لارتفاع القرار التصوير الزمانية المكانية للT خلايا البطانية المشبك المناعي كما ورد 19،20 وكذلك وصف هذا القانون. وكان من النتائج المركزي المستمدة من هذه مستو رواية الخلوية نموذج APC هو أن T ILPs خلية تعمل سواء في تعزيز الكشف حج الأولي والحفاظ على الإشارات اللاحقة. في الواقع، صفائف ILPs متعددة (التي استقرت والمستحقة في الرد المبدئي تدفق الكالسيوم) عرض تخصيب اليورانيوم في TCR والجزيئات توحي الإشارات النشطة مثل PKC-Q، ZAP-70، فسفوتيروزين وHS1. ولذلك، يبدو ILPs لتمثيل يعادل فيزيولوجي ثلاثي الأبعاد لالجزئي TCR-إشاراتمجموعات ينظر في نماذج طبقة ثنائية مستو. هذا النهج، وبالتالي، يكشف بحساسية / تقارير الديناميات الجزيئية والهندسة المعمارية (والضمنية النشاط الحيوي) لا يمكن كشفها خلاف ذلك.
الطريقة الموضحة في هذه الوثيقة يجب أن يكون مفيدا في سد الفجوة بين APC المهنية والصناعية نماذج الركيزة APC من أجل تعزيز قدرتنا على استجواب الآليات الأساسية من الاستجابات المناعية التكيفية. بينما هنا يتم التركيز على تفعيل CD4 + TH1 من نوع المستجيب / خلية الذاكرة، وهذا النهج الأساسي يمكن تعديلها بسهولة لدراسة مجموعة واسعة من أنواع الخلايا T وقسم الخدمات الزراعية، كما هو مبين أدناه.
