T lymfocyten een tak van het adaptieve immuunsysteem gekenmerkt door het vermogen om efficiënt te herkennen peptide-antigeen (Ag) gebonden aan major histocompatibility complex (MHC) moleculen door hun T-celreceptoren (TCR's) 1. Naïeve lymfocyten constitutief migreren en scannen 'professionele Ag presenterende cellen (APC's, bijvoorbeeld, dendritische cellen) in lymfeklieren, terwijl het geheugen / effector-T-cellen nodig hebben om effectief te overzien een zeer breed scala van APC's en potentieel doelwit cellen in perifere weefsels.
In de min na de eerste opname van verwante Ag op een APC, lymfocyten arresteren hun migratie en beginnen aan een gespecialiseerde intieme cel-cel-interface genaamd 'immunologische synaps' vormen (IS). Aanhoudende (dwz 30-60 min) IS contacten nodig zijn om te versterken en te ondersteunen signalering 2-7. Opkomende studies identificeren die binnen de IS, is de continue vorming en snelle remodeling van discrete sub-cellulaire signalering micro-clusters (dwz bevattende MHC / Ag-TCR, F-actine, adhesie en signaalmoleculen) de sterkte en de kwaliteit van de verkregen immuunreacties 07/02 bepalen. Echter, dynamische details en regulerend mechanisme van dit proces onvolledig begrepen 8,9. Dit vloeit grotendeels voort uit de technische uitdagingen in verband met onregelmatige topologieën van APC oppervlakken en slecht gecontroleerde oriëntatie van de cel-cel interactie vliegtuigen, kwesties die diep beperken de vereiste spatiotemporele imaging benaderingen 8-10 (Figure1A).
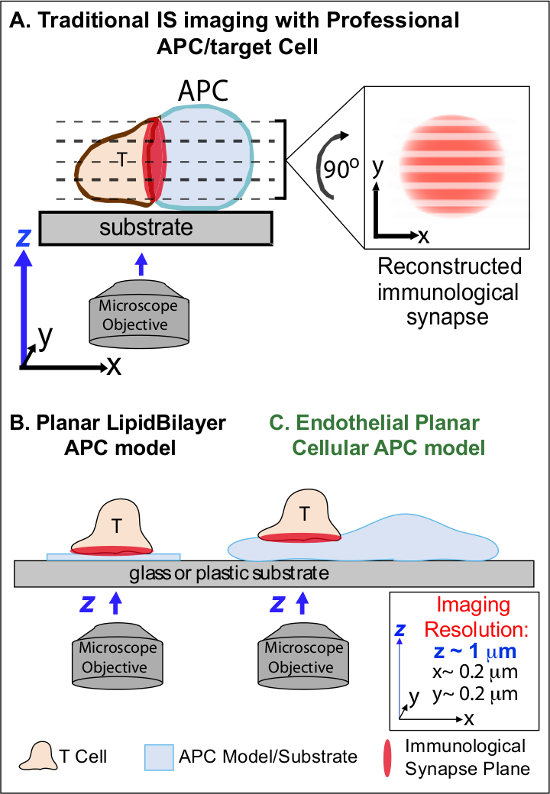
Figuur 1. Een Fysiologische Planar Cell APC Model for Imaging Immunologische Synapse Dynamics. Het schema illustreert traditionele beeldvorming van immunologische synaps tussen een T-cel en een professio nal APC (A) en T-cel en een traditionele vlakke lipide dubbellaag APC model (B) in relatie tot deze nieuwe endotheliale vlakke APC model (C). Professionele APCs bieden fysiologische immunologische synapsen maar bieden weinig georiënteerde cel-interface (dat wil zeggen ten opzichte van de optimale xy beeldvlak; resolutie ~ 0,2 pm), die dramatisch compromissen ruimtelijke (z beeldvlak resolutie ~ 1 micrometer) en tijd (dat wil zeggen, vanwege de noodzaak om herhaaldelijk gescand door alle z beeldvlakken) oplossing van beeldvorming. Bilaag modellen hebben een vlakke topologie die optimale spatiotemporele beeldvorming resolutie voorziet, maar ook sterk vereenvoudigd, niet-fysiologische en rigide. Deze endotheelcellen model combineert de vlakke topologie van lipidendubbellagen met de fysiologische substraat van een klassieke APC om een optimale ruimtelijke en temporele beeldvorming resolutie leveren in een fysiologische omgeving.m / files / ftp_upload / 53.288 / 53288fig1large.jpg "target =" _ blank "> Klik hier om een grotere versie van deze figuur te bekijken.
Vorige werk is gedeeltelijk omzeild deze obstakels door het ontwikkelen vlakke ondergrond modellen (dwz lipidendubbellagen en antilichaam-gecoate oppervlakken) die optimale spatiotemporele resolutie (dwz bieden, via de vaststelling van de T-cel activatie oppervlak in een enkel plan dat parallel aan de optimale xy beeldvorming vliegtuig) 11-15 (Figuur 1B). Deze modellen hebben belangrijke inzichten vergemakkelijkt in de subcellulaire / moleculaire dynamica dat antigene signalering in T-cellen, met inbegrip van de ontdekking van dynamische actine / TCR signalering micro-clusters 7,11-14 controleren. Echter, dergelijke modellen inherent oversimplified, evenals stijve (evenwel de ontwikkeling / studie van 3-dimensionale topologische kenmerken) (Figuur 1B). Daarom blijft het onzeker hoe dergelijke bevindingen aan phy betreffensiologic cel-cel immune surveillance.
Hoewel nog weinig bestudeerd, vasculaire en lymfatische endotheelcellen zijn in opkomst als een grote (dwz, groter in aantal dan alle professionele APC's, door ~ 1000-voudig) perifere compartiment van 'semi-professioneel "APC's 16-18. Deze cellen brengen MHC-I, MHC-II en een veelheid aan co-stimulator moleculen (bijvoorbeeld CD40, LFA3, ICOSL, 4-1BB, OX40L, TL1A, PD-L1, maar niet CD80 en CD86) en strategisch gepositioneerd aan de bloed-weefsel interface waar ze dienen gespecialiseerde sentinel functies 16-18. Vorige studies aangetoond dat endotheelcellen effectief opnieuw stimuleren effector / geheugen, maar niet naïeve T-cellen 19-25. Aldus endotheelcellen waarschijnlijk unieke APC rollen in effector fase van adaptieve immuunreacties in de perifere weefsels, zoals lokale invloed op T-celactivering, differentiatie, geheugen en tolerantie 16,17,26. Critisch, wanneer gekweekt in vitro endotheliale cellen vormen vrijwel vlakke celoppervlakken en gemakkelijk te transfecteren (bijvoorbeeld met fluorescente proteïne reporters). Deze functies zijn ideaal voor hoge spatiotemporele resolutie beeldvorming van topologische dynamiek in cel-cel interacties 19,27. Zo endotheelcellen kunnen dienen als een fysiologische "vlakke cellulaire APC-model duidelijk geschikt voor het onderzoek van de subcellulaire / moleculaire remodeling mechanismen antigeenherkenning drijven en te reguleren responsen (figuur 1C) 19,20.
Eerder vastgestelde complementaire beeldvormingstechnieken (waaronder transfectie van endotheel cellen met fluorescent eiwit makers van de plasmamembraan en cytosol) voor het bestuderen van de details van leukocyt-endotheel interactie tijdens hechting en transendotheliale migratie 27, bleek dat leukocyten actief sonde het oppervlak van het endotheel door dynamische inbrengen van eend terugtrekken van sub-micron-schaal, actine-rijke cilindrische uitsteeksels (~ 200-1000 nm in diameter en diepte) genoemd invadosome-achtige uitsteeksels (dat wil zeggen, 'POPs') 27,28. Deze beeldvormingstechnieken zijn verder uitgebreid met de creatie protocollen te profiteren van endotheliale APC functie om de eerste werkwijzen voor spatiotemporele hoge resolutie afbeelding van de T-cel-endotheliale immunologische synaps gerapporteerde 19,20 ontwikkelen en verder hierin beschreven. Een centrale bevinding afgeleid van deze nieuwe vlakke cellulaire APC model is dat T-cel POPs functioneren, zowel in het bevorderen van de initiële Ag detectie en bij het instandhouden van de daaropvolgende signalering. Inderdaad, arrays van meerdere POPs (die werden gestabiliseerd en opgebouwd in reactie op calciumflux initiële) tonen verrijking in TCR en moleculen die wijzen op actieve signalering zoals PKC-Q, ZAP-70, fosfotyrosine en HS1. Daarom POPs lijken een driedimensionale fysiologische equivalent aan het TCR-signalering micro vertegenwoordigenclusters gezien op vlakke dubbellaag modellen. Deze aanpak, dus onthult gevoelig / rapporten moleculaire en architecturale (en impliciete biomechanische) dynamiek anders niet aantoonbaar.
De hierin beschreven werkwijze moet nuttig zijn om de kloof tussen professionele APC en kunstmatige APC substraat modellen om ons vermogen om fundamentele mechanismen van adaptieve immuunreacties ondervragen verbeteren zijn. Hoewel hier de nadruk op de activatie van CD4 + Th1-type effector / geheugencel kan deze basisaanpak gemakkelijk worden aangepast aan een breed scala van T-cel soorten en Ags bestuderen, zoals hieronder besproken.
