T-Lymphozyten sind ein Zweig des adaptiven Immunsystems gekennzeichnet durch die Fähigkeit, Peptidantigen (Ag) effizient zu erkennen Haupthistokompatibilitätskomplex (MHC) gebundenen Moleküle durch ihre T-Zell-Rezeptoren (TCRs) 1. Naive Lymphozyten konstitutiv migrieren und scannen Berufs Ag-präsentierende Zellen "(APC; zB dendritische Zellen) innerhalb von Lymphknoten, während Memory / Effektor-T-Zellen benötigen, um eine extrem breite Palette von APCs und potentiellen Zielzellen im peripheren Gewebe effektiv zu überwachen.
Im min nach dem erstmaligen Ansatz des verwandten Ag auf einer APC, Lymphozyten zu verhaften ihre Migration und beginnen, eine spezialisierte intime Zell-Zell-Schnittstelle genannt 'immunologischen Synapse' zu bilden (IS). Anhaltende (dh 30-60 min) Kontakte sind erforderlich, um zu verstärken und zu erhalten Signalisierungs 2-7. Schwellen Studien identifizieren, die innerhalb der IS, ist es die kontinuierliche und schnelle Bildung remodeling diskreter subzellulären Signalisierungsmikrocluster (dh enthaltend MHC / Ag-TCR, F-Actin, Adhäsion und Signalmolekülen), die die Stärke und Qualität der resultierenden Immunantworten 2-7 zu bestimmen. Allerdings sind dynamische Details und regulatorischen Mechanismus dieses Prozesses unvollständig verstanden 8,9. Dies ergibt sich im Wesentlichen aus technischen Herausforderungen mit unregelmäßigen Topologien von APC Oberflächen und schlecht kontrollierte Ausrichtung der Zell-Zell-Interaktion Flugzeuge, Fragen, die zutiefst begrenzen die erforderliche Raumzeit-Bildgebung assoziiert nähert 8-10 (Abbildung 1a).
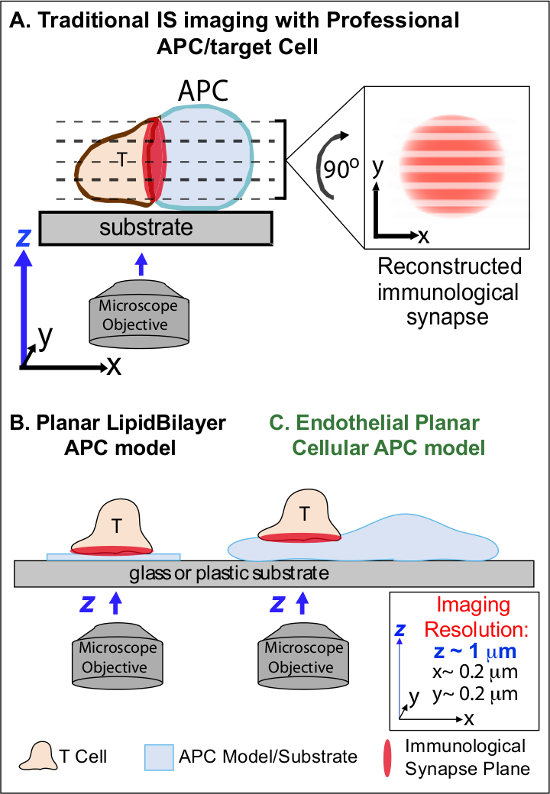
Abbildung 1. Ein Physiologische Planar Handy APC-Modell für Imaging immunologischen Synapse Dynamics. Die Abbildung zeigt, traditionellen Bildgebung von immunologischen Synapse zwischen einer T-Zelle und einer professio nal APC (A) und T-Zell-und ein traditionelles planaren Lipid-Doppelschicht APC-Modell (B) im Vergleich zu diesem Roman Endothelzellen planar APC-Modell (C). Professionelle APCs bereitzustellen physiologischen immunologischen Synapsen bieten aber schlecht orientierte Zell-Zell-Schnittstelle (dh in Bezug auf die optimale xy Abbildungsebene; Auflösung ~ 0,2 & mgr; m), die dramatisch beeinträchtigt räumliche (z Abbildungsebene Auflösung ~ 1 & mgr; m) und zeitliche (dh aufgrund der Notwendigkeit, immer wieder zu scannen durch alle z Abbildungsebenen) Auflösung der Bildgebung. Doppelschicht-Modelle haben eine planare Topologie, die eine optimale Raumzeit-Bildauflösung bietet, werden aber auch in stark vereinfachter, nicht-physiologischen und starr. Diese Endothelzellen-Modell vereint die planare Topologie der Lipid-Doppelschichten mit dem physiologischen Substrat eines klassischen APC, um eine optimale räumliche und zeitliche Bildauflösung in einer physiologischen Umgebung zu liefern.m / files / ftp_upload / 53288 / 53288fig1large.jpg "target =" _ blank "> Bitte klicken Sie hier, um eine größere Version dieser Figur zu sehen.
Frühere Arbeiten haben zum Teil diese Hindernisse durch Entwicklung planaren Substrat Modellen (dh Lipiddoppelschichten und Antikörper-beschichteten Oberflächen), welche optimale Raum-Zeit-Auflösung (dh bereitzustellen umgangen über Fixierung des T-Zell-Aktivierungsfläche in einem einzigen Plan, die parallel zu der optimalen xy Bildgebung Ebene) 11-15 (1B). Diese Modelle haben wichtige Einblicke in die subzelluläre / Molekulardynamik, die Antigen-Signalisierung in T-Zellen, einschließlich der Entdeckung der dynamischen Aktin / TCR Signalisierung Mikro-Clustern 7,11-14 Kontrolle erleichtert. Allerdings sind solche Modelle von Natur aus stark vereinfacht, sowie starre (Ausschluss der Entwicklung / Untersuchung der 3-dimensionale topologische Merkmale) (1B). Daher bleibt es unsicher, wie man solche Feststellungen phy beziehensiologic Zell-Zell-Immunüberwachung.
Obwohl noch under werden vaskulären und lymphatischen Endothelzellen als großen Schwellen (dh mehr in den Zahlen als alle professionellen APCs, die von ~ 1.000-fach) Peripheriefach 'semi-professionellen "APCs 16-18. Diese Zellen exprimieren MHC-I- MHC-II- und einer Vielzahl von Co-Stimulator Moleküle (zB CD40, LFA3, ICOSL, 4-1BB, OX40L, TL1A, PD-L1, nicht aber CD80 und CD86) und sind strategisch an der Blut-Gewebe-Grenzfläche, wo sie dienen spezialisierte Sentinel-Funktionen 16-18 positioniert. Frühere Studien zeigten, dass Endothelzellen wirksam neu zu stimulieren Effektor / Memory, aber nicht naiv, T-Zellen 19-25. Somit sind Endothelzellen wahrscheinlich einzigartig APC Rollen in Effektorphase adaptive Immunantworten in den peripheren Geweben, wie etwa lokale Einfluss auf T-Zellaktivierung, Differenzierung, Speicher und Toleranz 16,17,26 spielen. Crimatisch, wenn sie in vitro gezüchtet, Endothelzellen bilden praktisch planaren Zelloberflächen und leicht transfektierbare (beispielsweise mit fluoreszierenden Reporterproteine). Diese Eigenschaften sind ideal für hohe Raum-Zeit-auflösende Bildgebung der topologischen Dynamik während der Zell-Zell-Wechselwirkungen 19,27. So Endothelzellen als physiologische 'planaren Zell APC-Modell ausgesprochen geeignet für die Untersuchung der subzellulären / molekulare Umbaumechanismen, die Antigen-Erkennung zu fahren und zu regulieren Antworten (1C) 19,20 dienen könnte.
Vorher festgelegten komplementären Bildgebungstechniken (einschließlich Transfektion der Endothelzellen mit fluoreszierenden Protein Trägern der Plasmamembran und Cytosol) zur Untersuchung der Details der Leukozyten-Endothel-Interaktion während der Adhäsion und transendotheliale Migration 27, zeigte, daß Leukozyten die Oberfläche des Endothels Sonde aktiv durch dynamische Insertion eind Einfahren der Submikron-Maßstab, Actin-reiche zylindrischen Vorsprünge (~ 200-1000 nm Durchmesser und Tiefe) bezeichnet invadosome artigen Vorsprüngen (dh "ILPs ') 27,28. Diese bildgebenden Ansätze wurden weitere mit der Erstellung von Protokollen erweitert, um die Vorteile der endothelialen APC-Funktion, um die ersten Methoden zur hohen Raum-Zeit-auflösende Bildgebung des T-Zell-endothelialen immunologischen Synapse wie berichtet 19,20 entwickeln und weiter zu beschreiben hier. Eine zentrale Erkenntnis aus diesem neuartigen planaren Zell APC-Modell abgeleitet ist, dass T-Zell-ILPs funktionieren sowohl bei der Förderung der anfänglichen Ag Nachweis und bei der Aufrechterhaltung nachfolgende Signalisierung. Tatsächlich Arrays von mehreren ILPs (die stabilisiert und als Reaktion auf aufgelaufene Calciumfluss initial wurden) zeigen die Anreicherung in TCR und Molekülen andeutend aktives Melde wie PKC-Q, ZAP-70, Phosphotyrosin und HS1. Daher ILPs scheinen eine dreidimensionale physiologischen äquivalent der TCR-Signalgebung Mikro darstellenCluster in planare Doppelschicht-Modellen gesehen. Dieser Ansatz, so zeigt sensibel / Berichte molekularen und architektonischen (und stillschweigende biomechanischen) Dynamik sonst nicht nachweisbar.
Das hier beschriebene Verfahren ist nützlich, um die Lücke zwischen professionellen APC und künstliche APC-Substrat-Modelle, um unsere Fähigkeit, grundlegende Mechanismen der adaptiven Immunantwort zu verhören zu verbessern. Während hier der Schwerpunkt auf der Aktivierung von CD4 + Th1-Typ-Effektor / Speicherzelle kann dieser Ansatz leicht modifiziert werden, um eine breite Palette von T-Zell-Typen und Ags zu studieren, wie unten diskutiert.
