Groei analyse is afhankelijk van een set van tools die vaak worden gebruikt door planten wetenschappers om genotype bepaald groei verschillen en / of fenotypische reacties op milieu-factoren te beschrijven. Ze beschikken over de grootte en het gewicht metingen van de hele plant of een orgaan en berekeningen van groeicijfers om de onderliggende mechanismen van de groei te verkennen. Orgaangroei wordt bepaald door celdeling en expansie op cellulair niveau. Daarmee ook de kwantificering van deze twee processen in groeianalyses is essentieel voor het begrijpen van verschillen in groei 1 whole-orgaan. Bijgevolg is het cruciaal om een geschikte methodologie celgroei parameters die relatief eenvoudig te gebruiken door niet-gespecialiseerde laboratoria te bepalen.
Kinematische analyse reeds ingevoerd als een benadering die een krachtige kader voor de ontwikkeling van orgaangroei modellen 2. De techniek is geoptimaliseerd voor lineaire systemen,zoals Arabidopsis thaliana wortels en bladeren monocotyle, maar ook voor niet-lineaire systemen, zoals tweezaadlobbige bladeren 3. Tegenwoordig wordt deze methode steeds vaker gebruikt om te onderzoeken hoe genetische, hormonale, ontwikkelings- en milieufactoren invloed celdeling en groei in diverse organen (tabel 1). Bovendien biedt een raamwerk om cellulaire processen te verbinden met de onderliggende biochemische, moleculaire en fysiologische voorschriften (tabel 2), hoewel beperkingen door orgaangrootte en ruimtelijke organisatie kan worden opgelegd technieken die grotere hoeveelheden plantenmateriaal vereisen (bijv metaboliet metingen, proteomics, etc.).
Eenzaadlobbige bladeren, zoals maïs (Zea mays) bladeren vertegenwoordigen lineaire systemen waarbij cellen zich van de basis van het blad naar de punt, achtereenvolgens door het meristeem en elongatiezone de rijpe bereikenzone. Dit maakt het een ideaal modelsysteem voor kwantitatieve studies van de ruimtelijke patronen van de groei 4. Bovendien, maïs bladeren hebben grote groei zones (meristeem en rek zone verspreid over enkele centimeters 5) en bieden mogelijkheden om studies bij andere organisatieniveaus. Dit maakt het mogelijk voor het onderzoek naar de (vermeende) regulerende mechanismen die celdeling en expansie, gekwantificeerd door kinematische analyse door middel van een reeks van moleculaire technieken, fysiologische metingen, en celbiologie benaderingen (tabel 2).
Hier bieden we een protocol voor het uitvoeren van een kinematische analyse in eenzaadlobbige bladeren. Eerst wordt uitgelegd hoe een goede analyse van zowel celdeling en elongatie voeren als functie van positie langs de vleugelas en hoe kinematische parameters berekenen. Ten tweede hebben wij laten ook zien hoe deze kan worden gebruikt als basis voor de steekproef. Hier bespreken we twee gevallen: high-resolution sampling eend gerichte steekproeven, waardoor betere interpretatie van gegevens en de besparing van tijd / geld, respectievelijk.
Tabel 1. Overzicht van kinematische analysemethoden voor kwantificeren van celdeling en groei in diverse organen.
| orgaan | referentie |
| eenzaadlobbige bladeren | 16, 20, 21, 22 |
| Root tips | 2, 23, 24, 25, 26, 27, 28, 29 |
| tweezaadlobbige bladeren | 21, 30, 31 |
| schieten topmeristeem | 32 |
Tabel 1. Overzicht van kinematische analysemethoden voor kwantificeren van celdeling en groei in diverse organen.
<p class="jove_content" fo:keep-together.within-page = "1">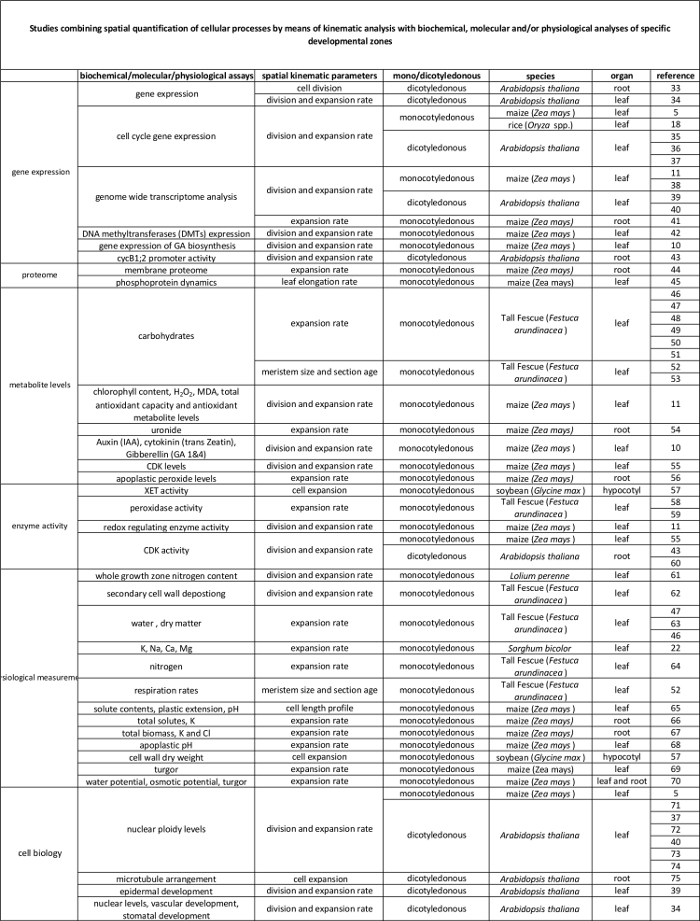
Tabel 2. Verband tussen cellulaire processen gekwantificeerd door de kinematische analyse om de regulering op moleculair niveau. Verwijzingen naar verscheidene studies die de kwantificering van cellulaire processen om resultaten van biochemische en moleculaire assays verschillende soorten en organen. Xyloglucan endotransglucosylase (XET), malondialdehyde (MDA), cycline-afhankelijke kinasen (CDK). Klik hier om een grotere versie van deze tabel te bekijken.
