Maïs blad een ideaal model voor de vorming van ontwikkelings- gebieden te bestuderen bij morfogenese, aangezien het een duidelijke grens tussen het mes en schede die vatbaar is voor genetische dissectie (Figuur 1A). Tijdens de vroege stadia van bladontwikkeling, een lineaire band kleinere cellen, de preligule band (PLB), verdeelt het blad primordium in vooraf blad en pre-mantel domeinen. Een pony-achtig tongetje en driehoekige oorschelpen ontwikkelen van de PLB (Figuur 1 A, C, D). Genetische screens hebben geïdentificeerd mutaties die het mes-schede grens verstoren. Bijvoorbeeld recessieve liguleless1 (LG1) mutaties te verwijderen en de ligule oorschelpen 1, 2, 3, 4 (figuur 1B). In situ hybridisatie toonde dat LG1 transcript ophoopt op de PLB en opkomende tongetje, waardoor het een uitstekende marker voor tongetje ontwikkeling 5, 6 (figuur 1E).
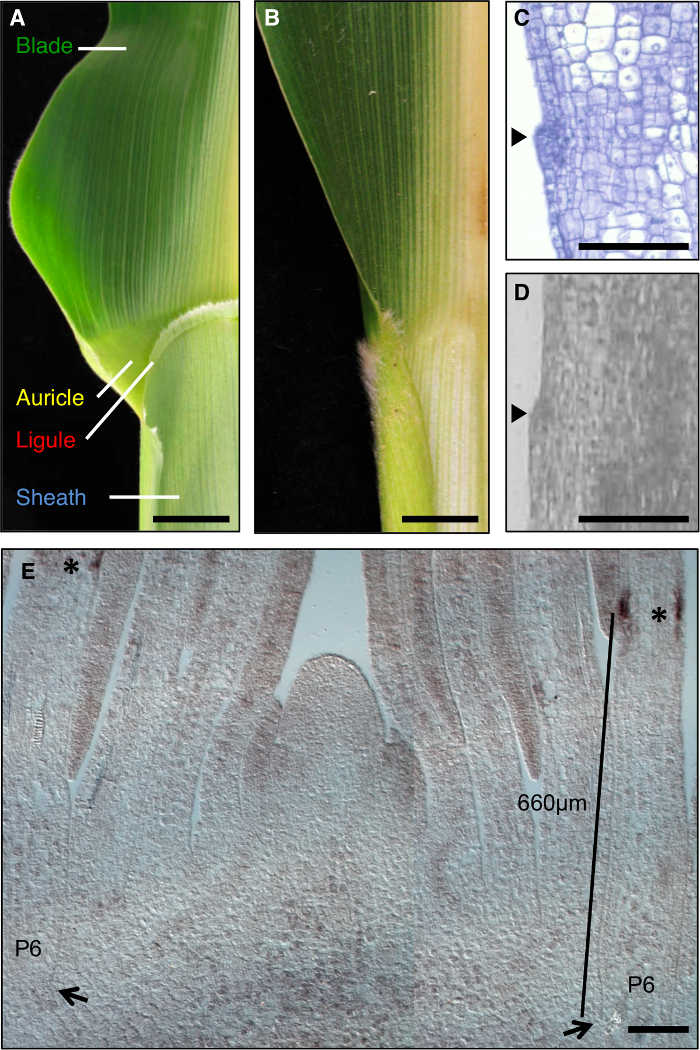
Figuur 1: Wild-type en liguleless1-R maïs bladeren. (A) Blade-omhulsel grensgebied van volwassen wild-type blad met tongetje en oorschelp structuren. (B) Blade-omhulsel grensgebied van volwassen liguleless1-R blad met afwezigheid van tongetje en oorschelp structuren. Bladeren in A en B zijn gesneden in de helft langs de hoofdnerf. (C) langsdoorsnede door wild-type blad primordium. Monster is verwerkt en gekleurd voor histologische analyse. Het initiërende tongetje blijkt als een bult uitsteekt uit het vlak van het blad (pijlpunt). (D) Longitudinal sekteion door middel van wild-type blad primordium. Monster is verwerkt LM zoals beschreven in de tekst. Arrowhead geeft initiëren tongetje. (E) LG1 in situ hybridisatie van shoot apex laterale langsdoorsnede. Sterretjes geven LG1 transcript ophoping aan de PLB van de P6 blad primordium. Pijlen geven voet van P6 primordium. Bar geeft meting van de basis van de primordium de PLB. Schaalbalk in A en B = 20 mm. Schaal bars in CE = 100 urn. Dit cijfer is gewijzigd ten opzichte van referentie-6 (Copyright American Society of Plant Biologen). Klik hier om een grotere versie van deze figuur te bekijken.
In deze studie werd LM RNA-Seq gebruikt om een reeks van genen die differentieel tot expressie (DE) geeft bij het mes omhulsel grens ten opzichte van andere delen van het blad primordium en ide ntify genen die in DE LG1-R mutanten ten opzichte van wild-type broers en zusters. LM RNA-Seq is een werkwijze voor het kwantificeren transcript accumulatie in specifieke cellen of cellulaire domeinen 7. LM systemen combineren een laser en een microscoop met een digitale camera. Doorsnede weefsel wordt aangebracht op objectglaasjes en bekeken door de microscoop. De LM-software bevat meestal tekentools waarmee de gebruiker kan worden geselecteerd gebied te schetsen voor microdissectie. De laser snijdt langs de lijn, en het gekozen weefsel wordt gekatapulteerd van de glijbaan en in een buis boven de dia opgeschort. LM kan de gebruiker nauwkeurige domeinen, inclusief specifieke cellagen en zelfs individuele cellen 8, 9 microdissect. RNA kan dan worden geëxtraheerd uit het weefsel gemicrodissecteerde. Vervolgens worden de RNA-component gebruikt Seq volgende generatie sequencing van cDNA-bibliotheken gegenereerd uit het geëxtraheerde RNA sequentie 10,= "xref"> 11.
Belangrijkste voordelen van LM RNA-seq zijn de mogelijkheid om transcript ophoping kwantificeren welbepaalde domeinen en het vermogen om de hele transcriptoom gelijktijdig 7 profileren. De techniek is bijzonder geschikt voor probing vroege ontwikkelingsgebeurtenissen wanneer het interessegebied vaak microscopisch. Eerdere studies hebben gebruikt LM gecombineerd met microarray technologie ontwikkelingsprocessen te bestuderen in planten 9, 12, 13. RNA-Seq heeft het voordeel kwantificeren transcripten over een breed dynamisch bereik, waaronder lage-genen tot expressie, en voorafgaande sequentie-informatie is niet vereist 10, 11. Bovendien LM RNA-Seq heeft het potentieel om ontwikkelings belangrijk genen die kunnen ontbreken in mutagenese schermen door genetische redundantie of letaliteit van het verlies-van- markerenfunctie mutant.
Ontwikkelings belangrijk genen, zoals smalle sheath1 (NS1) pt bekervormige cotyledon2 (cuc2), vaak specifieke expressiepatronen van slechts één of enkele cellen 17, 18, 19, 20. Velen zijn uitgedrukt alleen tijdens de vroege ontwikkelingsstadia en niet in het rijpe orgaan. Wanneer gehele organen of grote domeinen worden geanalyseerd, worden deze celspecifieke transcripten verdund en kunnen niet meer conventionele analyses worden gedetecteerd. Door toe te staan analyses van precies gedefinieerde domeinen, LM RNA-Seq maakt deze weefsel-specifieke genen te identificeren en te kwantificeren.
Cruciaal voor het succes van de hier beschreven experimenten een grondige histologische analyse dat de selectie van de geschikte ontwikkelingsstadium en domein voor analyse geleid en nauwkeurige MeasureMent van celweefsel domeinen LM. Opdat equivalente domeinen bemonsterd over alle replicaten werd weefsel vanuit bladprimordia in hetzelfde ontwikkelingsstadium en gemicrodissecteerde domeinen opzichte van morfologische oriëntatiepunten gemeten zoals de opkomende ligule (figuur 2). Het is bekend dat sommige genen tot expressie gebracht in een gradiënt van de top tot de basis van het blad. Door het meten van nauwkeurige domeinen variëren door bemonstering vanaf verschillende plaatsen langs het blad proximaal-distale as werd minimaal gehouden (figuur 3A). Door microdissecting domeinen van dezelfde grootte, variatie door het differentiële verdunning van celspecifieke transcripten werd verlaagd (Figuur 3B). Zijdelingse langsliggers die de shoot apex gebruikt voor microdissections. Deze doorsneden loodrecht op de hoofdnerf marge as (Figuur 4). Met alleen secties die de SAM bevatten zorgt ervoor dat gelijkwaardige laterale regio's vanbladprimordia worden geanalyseerd.
In monsters verwerkt en deelbaar LM, de eerste morfologische tekenen van uitgroei tongetje een bult op de adaxiale kant door periclinale celdelingen in de adaxiale epidermis (figuur 1D, figuur 2). Er werd vastgesteld dat de opkomende ligule betrouwbaar kan worden geïdentificeerd plastochron 7 stadium bladprimordia. We waren geïnteresseerd in genen die tot expressie in het gehele tongetje regio, inclusief de nieuwe tongetje en de cellen onmiddellijk distaal dat de oorschelp vormen. Om te waarborgen dat gelijkwaardige weefsel selecties gemaakt, werd het tongetje bult als morfologisch landmark en 100 urn rechthoek gecentreerd op de ligule bult werd geselecteerd LM (figuur 2A, 2B). Equivalent sized rechthoeken van pre-blad en pre-mantel geselecteerd uit dezelfde bladprimordia.
Analyses van liguleless mutant planten presenteerde een andere challeNSE; LG1-R mutanten geen tongetje dus deze morfologische eigenschap kan niet worden gebruikt om het gebied te selecteren LM vormen. In plaats daarvan, het domein van LG1 transcript accumulatie in wildtype bladprimordia werd bepaald, en een gebied dat domein zou omvatten gedefinieerd. Deze voorlopige analyses werden uitgevoerd op zaailingen van dezelfde planten als werden gebruikt voor de uiteindelijke analyse, omdat eerder werk heeft aangetoond dat de ligging van de PLB afhankelijk van groeiomstandigheden. In situ hybridisatie aangegeven dat LG1 transcripten accumuleren in de PLB van P6 bladprimordia (figuur 1E). We selecteerden een domein 400-900 urn van de basis van de bladprimordia dat het domein van LG1 expressie (paarse bouwstenen, figuur 2A) omvat en gevangen deze equivalente gebieden van wildtype en LG1-R planten. De variatie in genetische achtergrond en groeiomstandigheden minimaliseren bij vergelijking transcript accumulatie in LG1-R en wild-type planten, scheiden families van mutanten en wild-type broers en zussen werden gebruikt.
