Die Maisblatt ist ein ideales Modell die Bildung von Entwicklungsfelder während der Morphogenese zu untersuchen, da es eine deutliche Grenze zwischen der Klinge und Hülle aufweist , die zu genetischen Dissektion (1A) zugänglich ist. Während der frühen Stadien der Blattentwicklung, einem linearen Band von kleineren Zellen, die preligule Band (PLB), unterteilt die Blatt primordium in vorge Klinge und pre-Mantel-Domänen. Ein saumartig ligule und Dreiecks Vorhöfe entwickeln sich aus der PLB (1A, C, D). Genetische Screens identifiziert Mutationen, die die Klingenscheide Grenze unterbrechen. Zum Beispiel, rezessiv liguleless1 (LG1) Mutationen löschen Sie die ligule und Vorhöfe 1, 2, 3, 4 (1B). In – situ – Hybridisierung zeigte , dass LG1 Transkript sammelt sich an der PLB und Schwellen ligule, es eine ausgezeichnete Marker für ligule Entwicklung 5, 6 (Abbildung 1E) zu machen.
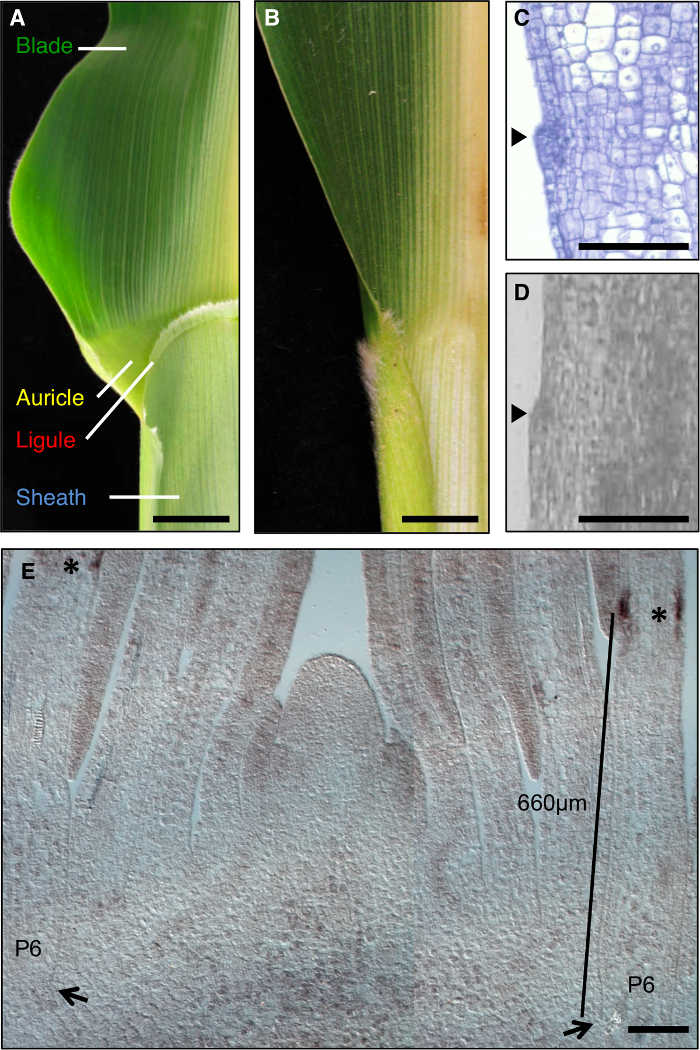
Abbildung 1: Wildtyp und liguleless1-R Maisblätter. (A) Blattscheide Randbereich des reifen Blatt Wildtyp zeigt ligule und Ohrmuschel Strukturen. (B) Blattscheide Grenzbereich von reifen liguleless1-R Blatt zeigt Abwesenheit von ligule und Ohrmuschel Strukturen. Blätter in der A und B wurden in zwei Hälften entlang der Mittelrippe geschnitten. (C) Längsschnitt durch Wildtyp – Blatt primordium. Probe wurde für die histologische Analyse verarbeitet und gefärbt. Der initiierende ligule ist offensichtlich, wie eine Unebenheit von der Ebene des Blattes vorsteht (Pfeilspitze). (D) Längs SekteIon durch Wildtyp-Blatt primordium. Probe wurde für LM verarbeitet, wie im Text beschrieben. Pfeilspitze zeigt ligule initiieren. (E) LG1 in – situ – Hybridisierung von Spross seitlichen Längsschnitt. Sternchen zeigen LG1 Transkriptakkumulation an der PLB des P6 Blatt primordium. Die Pfeile zeigen die Basis von P6 primordium. Balken zeigt Messung von der Basis des primordium zum PLB. Maßstabsbalken in A und B = 20 mm. Maßstabsbalken in CE = 100 & mgr; m. Diese Zahl wurde von Referenz 6 (Urheberrecht American Society of Plant Biologists) modifiziert. Bitte klicken Sie hier , um eine größere Version dieser Figur zu sehen.
In dieser Studie wurde LM RNA-Seq verwendet eine Reihe von Genen zu identifizieren, die differentiell exprimierte (DE) an der Messerscheide Grenze relativ zu anderen Teilen des Blatt primordium und ide ntify Gene , die DE in LG1-R – Mutanten im Vergleich zu Wildtyp – Geschwister sind. LM RNA-Seq ist ein Verfahren der 7 Transkriptakkumulation in spezifischen Zellen oder zellulären Domänen zu quantifizieren. LM-Systeme kombinieren einen Laser und ein Mikroskop mit einer Digitalkamera. Geteilter Gewebe wird auf Schlitten montiert und durch das Mikroskop betrachtet. Die LM-Software enthält in der Regel Zeichenwerkzeuge, die dem Benutzer ermöglichen, alle ausgewählten Region für die Mikrodissektion zu skizzieren. Die Laserschnitte entlang der Linie, und das ausgewählte Gewebe wird die Folie und in ein Rohr über dem Objektträger aufgehängt katapultiert. LM ermöglicht es dem Benutzer präzise Domains microdissect, einschließlich spezifischer Zellschichten und sogar einzelne Zellen 8, 9. RNA kann dann aus dem microdissected Gewebe extrahiert werden. Anschließend wird die RNA-Seq Komponente der nächsten Generation Sequenzierung Sequenzierung von cDNA – Bibliotheken erzeugt aus der extrahierten RNA 10 verwendet,= "xref"> 11.
Die wichtigsten Vorteile von LM RNA-seq sind die Fähigkeit , Transkriptakkumulation in genau definierten Bereichen und die Fähigkeit zum Profil des gesamten Transkriptom gleichzeitig 7 zu quantifizieren. Die Technik eignet sich besonders für frühe Entwicklungs Ereignisse Sondieren, wo die Region von Interesse ist oft mikroskopisch. Frühere Studien haben LM mit Microarray – Technologie in Kombination verwendet , um Entwicklungsprozesse in Pflanzen 9, 12, 13 studieren. RNA-Seq hat den Vorteil der Transkripte in einem breiten Dynamikbereich, einschließlich niedrig exprimierten Genen Quantifizieren und vor Sequenzinformation ist nicht erforderlich , 10, 11. Darüber hinaus hat LM RNA-Seq das Potenzial ihrer Entwicklung wichtige Gene hervorzuheben, die durch genetische Redundanz in Mutagenese-sehen werden oder zu Letalität der Verlust-of-Funktion Mutante.
Entwicklungs wichtige Gene, wie schmal sheath1 (NS1) und becherförmigen cotyledon2 (CUC2), haben oft spezifische Expressionsmuster von nur einem oder wenigen Zellen 17, 18, 19, 20. Viele von ihnen sind in der frühen Entwicklungsstadien und nicht in der reifen Organ nur zum Ausdruck gebracht. Wenn ganze Organe oder große Domänen analysiert werden, wobei diese zellspezifische Transkripte werden verdünnt und kann nicht in konventionelleren Analysen nachgewiesen werden. Durch Analysen von genau definierten Domänen ermöglicht, LM RNA-Seq ermöglicht diese gewebespezifischen Gene identifiziert und quantifiziert werden.
Entscheidende Faktoren für den Erfolg der hier beschriebenen Experimente wurden eine gründliche histologische Analyse, die Auswahl der geeigneten Entwicklungsstadium und Domäne für die Analyse geführt und präzise measurement von Zell-Gewebe-Domänen für LM. Um sicherzustellen , dass äquivalente Domänen für alle Wiederholungs abgetastet wurden, Gewebe wurde aus Blattprimordien auf der gleichen Entwicklungsstadium gesammelt und die microdissected Domänen wurden relativ gemessen zu morphologischen Orientierungspunkte wie die Schwellen ligule (Abbildung 2). Es ist bekannt, dass einige Gene in einem Gradienten von der Spitze bis zur Basis des Blattes exprimiert werden. Durch die Messung präzise Domänen Variation aufgrund der Abtastung von verschiedenen Stellen entlang des Blatt proximal-distalen Achse auf ein Minimum (3A) gehalten werden . Von Domänen der gleichen Größe microdissecting Variation aufgrund unterschiedlicher Verdünnung von zellspezifische Transkripte wurde ebenfalls reduziert werden (3B). Seitliche Längsschnitte des Spross wurden für alle Mikrodissektionen verwendet. Dies sind Bereiche , die senkrecht zur Mittelrippe-margin Achse (Abbildung 4). Mit nur Abschnitte, die die SAM umfassen wird sichergestellt, dass äquivalente Seitenbereiche vonBlattprimor analysiert.
In Proben verarbeitet und LM, der erste morphologischen Zeichen von ligule Auswuchs geschnitten ist eine Beule auf der oberseits aufgrund perikline Zellteilungen in den adaxial Epidermis (1D, 2). Es wurde festgestellt, dass die Schwellen ligule zuverlässig bei Plastochron 7 Stadium Blattanlagen identifiziert werden konnten. Wir waren daran interessiert, Gene in der gesamten Region ligule ausgedrückt, einschließlich der Schwellen ligule und die Zellen unmittelbar distal, die die Ohrmuschel bildet. Um sicherzustellen , dass Selektionen äquivalente Gewebe hergestellt wurden, wurde die ligule Bump als morphologische Wahrzeichen verwendet und eine 100 & mgr; m Rechteck auf der ligule Bump zentriert wurde LM (2A, 2B) ausgewählt. Equivalent große Rechtecke vorge Klinge und pre-Mantel wurden aus den gleichen Blattprimordien ausgewählt.
Die Analysen der liguleless mutierten Pflanzen präsentiert eine andere challenge; LG1-R – Mutanten bilden keine ligule, daher ist diese morphologische Funktion nicht verwendet werden, um die Region für LM auszuwählen. Stattdessen wurde die Domäne von LG1 Transkriptakkumulation in Wildtyp – Blattanlagen bestimmt, und eine Region , die diese Domäne wurde definiert umfassen würde. Diese vorläufigen Analysen wurden auf Sämlingen aus demselben Anpflanzung durchgeführt, wie für die endgültige Analyse verwendet wurden, da frühere Arbeiten haben gezeigt, dass der Ort des PLB variiert je nach Wachstumsbedingungen abhängig. In – situ – Hybridisierung angegeben , dass LG1 – Transkripte in der PLB von P6 Blattanlagen (Abbildung 1E) akkumulieren. Wir haben für Sie eine Domäne 400-900 & mgr; m von der Basis des Blattes primordia, die die Domäne von LG1 Ausdruck umfasste (lila Rechtecke, 2A) und erfasst diese äquivalenten Regionen von Wildtyp und LG1-R Pflanzen. Zur Minimierung der Variation in der genetischen Hintergrund und Wachstumsbedingungen bei einem Vergleich TRANSCRIPt Akkumulation in LG1-R und Wildtyp – Pflanzen, Aussortieren Familien von Mutanten und Wildtyp – Geschwister wurden verwendet.
