Figure 1 shows a schematic representation of the cell-seeding and CLC:CLF-1 incubation in protocol 1. Figure 2 shows a schematic representation of the cell-seeding and leu/pep and CLC:CLF-1 incubation in protocol 2. Figure 3 shows a schematic representation of the cell-seeding and CLC:CLF-1 incubation and stimulation in protocol 3. Figure 4 shows the sorLA-mediated CLC:CLF-1-dependent downregulation of CNTFRα. Figure 5 shows the sorLA-mediated endocytosis and lysosomal targeting of CNTFRα. Figure 6 shows the lower response to CLC:CLF-1 stimulation after CNTFRα downregulation.
Note that bands signifying CNTFRα-Myc have similar density at time zero, whereas downregulation of CNTFRα-Myc is demonstrated by the weaker band in sorLA transfectants after 5 h (Figure 4). Figure 5 shows that CNTFRα-Myc has accumulated in LAMP-1 positive vesicles of leu/pep treated cells. In contrast, no accumulation of CNTFRα-Myc is seen in cells not subjected to leu/pep treatment. This demonstrates that CNTFRα-Myc is sorted to lysosomes and degraded. As can be seen in Figure 6, the level of pSTAT3 is significantly reduced in pre-stimulated cells as compared to the level in cells that has not been exposed to CLC:CLF-1. This is in accordance with the observed sorLA-mediated downregulation of CNTFRα in the pre-stimulated cells (Figure 4).
Figure 1: Schematic representation of cell-seeding and CLC:CLF-1 incubation in protocol 1. Seed HEK293-CNTFRα-Myc and HEK293-CNTFRα-Myc/sorLA cells in two 4-well plates (0 and 5 h) and incubate cells with CLC:CLF-1 as indicated. Please click here to view a larger version of this figure.
Figure 2: Schematic representation of cell-seeding and leu/pep and CLC:CLF-1 incubation in protocol 2. Seed HEK293-CNTFRα-Myc/sorLA cells on cover slides in a 4-well plate and incubate cells with or without leu/pep and CLC:CLF-1 as indicated. Please click here to view a larger version of this figure.
Figure 3: Schematic representation of cell-seeding and CLC:CLF incubation and stimulation in protocol 3. Seed HEK293-sorLA cells in one 4-well plate and incubate cells with or without CLC:CLF-1 (pre-exp.) to downregulate the CNTFRα followed by CLC:CLF-1 stimulation (stim.) to initiate STAT3 phosphorylation as indicated. Please click here to view a larger version of this figure.
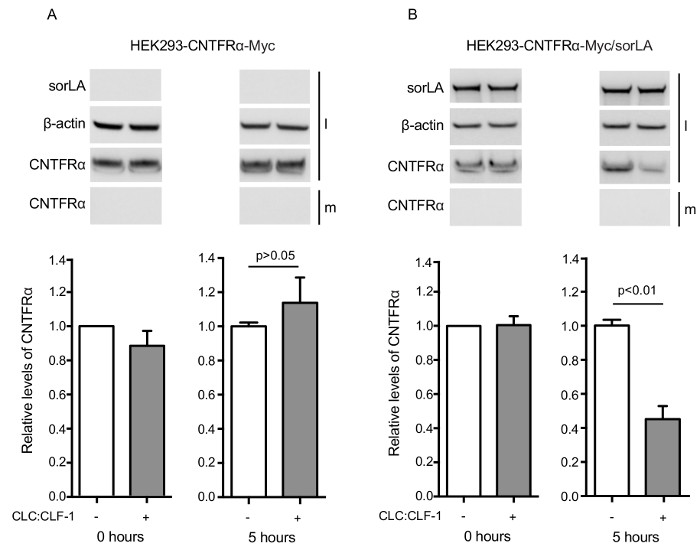
Figure 4: Results showing that SorLA mediates CLC:CLF-1-dependent downregulation of CNTFRα. (A) HEK293-CNTFRα-Myc and (B) HEK293-CNTFRα-Myc/sorLA cells were incubated in the absence (white columns) or presence (grey columns) of 10 nM CLC:CLF-1. After 0 and 5 h, the incubation was stopped and the content of CNTFRα-Myc in the medium (m) and in the cell lysates (l) was detected by Western blotting and quantified by densitometry. The upper panels show Western blot results from representative experiments and the lower panels show the detected levels of CNTFRα-Myc found in the cell lysates. The levels are shown relative to the CNTFRα-Myc level in the single and double transfectants at 0 h. Each column represents mean ±SEM (n = 3). p-values calculated using t-test. Reproduced after original figure13. Please click here to view a larger version of this figure.
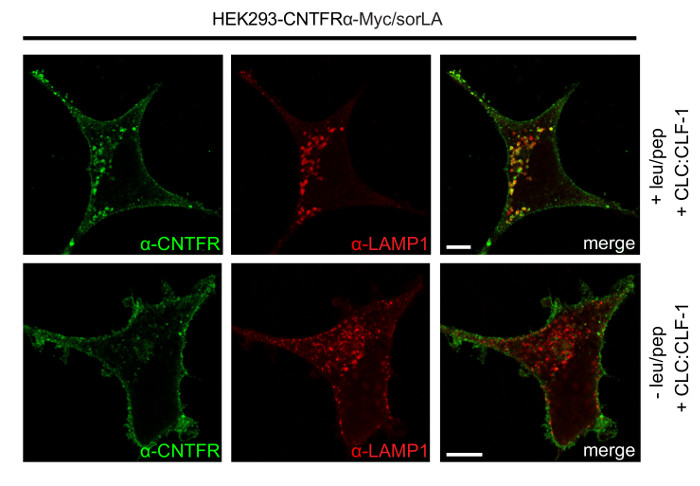
Figure 5: Results showing that SorLA mediates endocytosis and lysosomal targeting of CNTFRα. HEK293-CNTFRα-Myc/sorLA cells were treated with or without leu/pep, incubated with 10 nM CLC:CLF-1 for 5 h, fixed, and finally stained using anti-CNTFRα and anti-LAMP-1 antibodies as described in protocol 2. Scale bars: 5 μm. Reproduced after original figure13. Please click here to view a larger version of this figure.
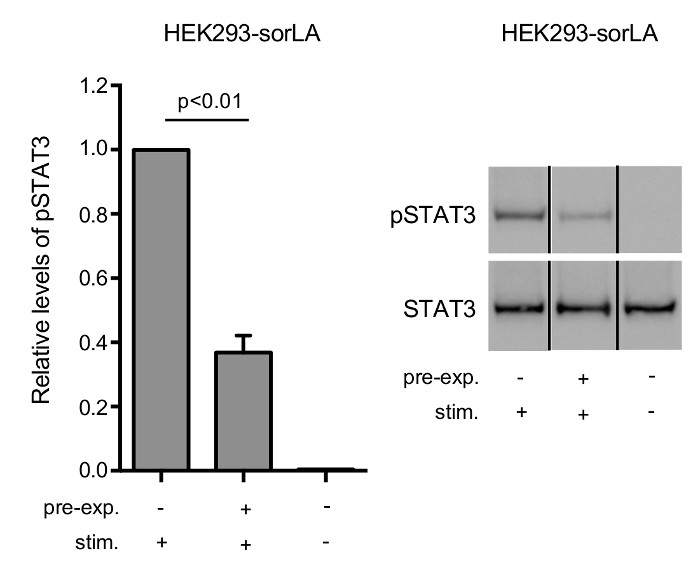
Figure 6: SorLA-mediated downregulation of CNTFRα is accompanied by a lowered cellular response to CLC:CLF-1 stimulation.
HEK293-sorLA cells were pre-incubated in the absence or presence of 10 nM CLC:CLF-1 for 5 h (pre-exp.), starved in blank medium for 90 min, and stimulated with 5 nM CLC:CLF-1 for 15 min (stim.). (left) The columns show the relative levels of pSTAT3 in the cells. Each column represents mean ±SEM (n = 3) relative to the pSTAT3 level in cells preincubated in the absence of CLC:CLF-1 but stimulated with CLC:CLF-1. p-value calculated using t-test. (right) The Western blot shows the response of pSTAT3 and STAT3 obtained in a representative experiment. Reproduced after original figure13. Please click here to view a larger version of this figure.
