Candida-arter er commensal sopp som kolonisere tarm og urogenitale områder av alle mennesker. Under tilstander med immunsvikt, så som det som forekommer med prematur fødsel eller immunsuppressive effekter av behandlinger for kreft, kan Candida-arter bli opportunistiske patogener. Av de Candida-arter, er Candida albicans den mest utbredte sopp kolonisator og forårsaker de fleste av invasive soppinfeksjoner. Andre Candida-arter som C. glabrata, C. parapsilosis, C. tropicalis og C. kruseii også forårsake alvorlige infeksjoner hos immunkompromitterte pasienter, med noen oppviser iboende motstand mot brukte anti-fungal antibiotika slik som fluconazol og amfotericin B. Derfor infeksjoner med noen av disse artene blir observert oftere, spesielt hos pasienter som behandles profylaktisk med antisoppmidler. Selv med riktig og rettidig enNTI-soppbehandling, invasive Candida-infeksjoner fortsette å være assosiert med signifikant morbiditet og mortalitet 1. På grunn av betydningen av Candida-arter i menneskers helse, er det et behov for lett tilgjengelige molekylære verktøy som tillater studiet og klarlegging av deres patogenese mekanismer.
Et viktig verktøy som lar forskere å visualisere og kvantifisere mikrobielle celler og proteiner som de uttrykker er FP fusion teknologi. Polymerase kjedereaksjon (PCR) -mediert gen modifikasjon, slik det er beskrevet i denne artikkelen, tillater bygging av fusjoner mellom FP sekvenser og en Candida-protein-kodende sekvens av interesse ved dens genomiske locus. Stabil integrering av konstruksjonen forenkler analysen av protein ekspresjon, så vel som proteinlokaliserings dynamikk. Plasmider inneholdende sekvenser som FP, optimalisert for ekspresjon i Candida albicans, og som kan brukes i PCR-mediert gene modifikasjon strategi, er tidligere bygget 2, 3, 4, 5. Plasmider som inneholder FP transformasjon "kassetter": et FP-sekvens bundet til et ernæringsmessig markørgen som muliggjør transformasjon av C. albicans og C. parapsilosis 2, 3, 4, 5, 6, 7. For tiden tilgjengelige plasmider inneholder en rekke valgbare markørgener ernærings (URA3, HIS1, ARG4) for omforming av auxotrofe stammer, så vel som en dominant medikamentresistensmarkør (Nat1), som muliggjør transformasjon av kliniske stammer mangler auxotrophies. I tillegg plasmider inneholder muligheter for opptil fire forskjellige FP sekvenser (grønn [GFP], yellow [YFP], cyan [CFP] og kirsebær [mCherry]) og enten en ADH1 termineringssekvens for bygging av carboxyenden proteinfusjoner, eller en promotorsekvens for bygging av amino-terminale proteinfusjoner. Primerne er utformet med homologi med plasmid-DNA som omgir FP kassetten. I tillegg primerne inneholder også 5'-forlengelse sekvenser som bærer homologi til gjærgenet av interesse å bli merket, noe som letter integrering av kassetten i det genomiske lokuset via homolog rekombinasjon (figur 1). Genspesifikke FP-kassetter er generert ved PCR og deretter transformert inn i Candida-celler som er gjort kompetente for opptak av DNA ved behandling med litium-acetat.
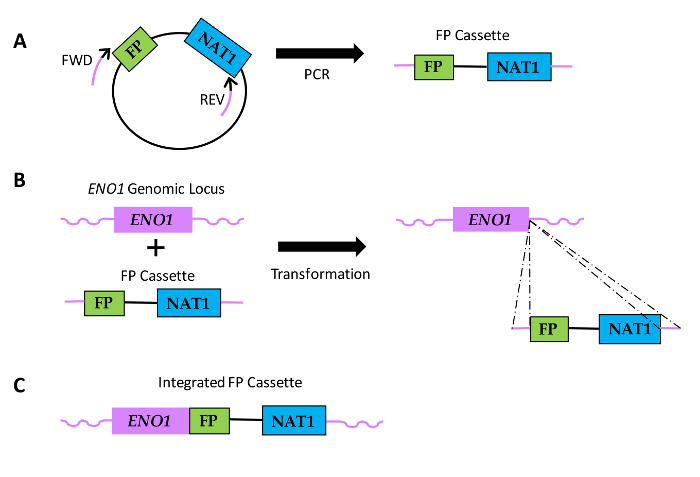
Figur 1: Diagram av hvordan FP sekvens fusjoner blir generert i Candida-arter. (A) Plasmid-DNA inklues en FP sekvens og en sekvens som koder nourseothricin motstand (Nat1). Relative steder av Forward (FWD) og revers (REV) primere er vist med svarte deler av primere som indikerer regionen av homologi til plasmidet sekvens og lilla deler betegner genet spesifikke homologi region eller primer extension. (B) FP kassetter er forvandlet til Candida og integrere i ENO1 genomiske locus via homolog rekombinasjon (stiplede linjer). (C) Resulterer FP fusjon sekvens på 3'end av ENO1. Klikk her for å se en større versjon av dette tallet.
Heri presenterer vi et eksempel på proteinfusjon (Eno1-FP) konstruksjoner i Candida-arter. Vi bruker tagging plasmider som inneholder Nat1 transformasjon markørgenet sammen med sekvenser som koder GFP, YFP, ellermCherry (figur 2). Disse plasmidene brukes sammen med primere i PCR for å generere genspesifikke kassetter som lette fusjon av fps til 3'-enden av ENO1, noe som resulterer i ekspresjon av Eno1 kondensert til fps ved sin karboksy-terminus.
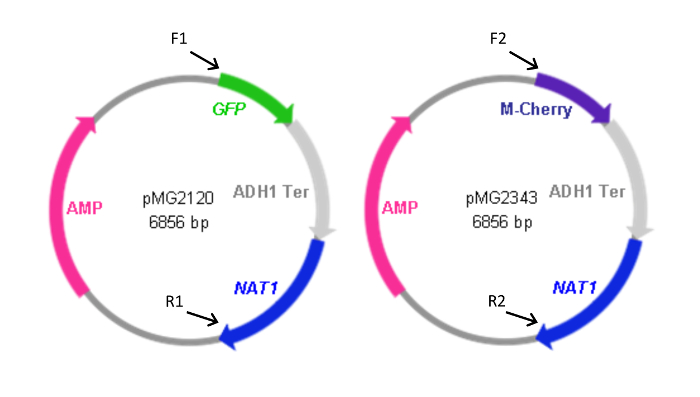
Figur 2: Kart over FP kassett som inneholder plasmider. Forover (F) og omvendt (R) primere som brukes for å generere kassettene fra plasmidene er angitt sammen med den relative plasseringen av deres homologi til plasmidene. Primer sekvenser er som angitt i tabell 1. F1 og R1 ble også brukt for å generere pYFP- Nat1 kassetten. Plasmidet inneholdende YFP- Nat1 kassetten (pMG2263) er identisk med pMG2120 med unntak av YFP i stedet for GFP-sekvensen. Kassett størrelser: GFP-Nat1, 3,7 KBP; mCherry- Nat1, 3,2 kbp; YFP- Nat1, tre.7 kbp. Dette tallet har blitt forandret fra Gerami-Nejad, et al. 4 Klikk her for å se en større versjon av dette tallet.
