We describe a flexible and efficient method to generate discrete spheroids using cell-repellent plates and spheroid formation media supplemented with MC. Under the appropriate conditions of MC and serum, individual cells settle and adhere together at the center of the well to form spheroids with minimal adherence to the well bottom. Using this protocol, spheroids were generated from a variety of cell lines (Figure 2B). Titration of MC and serum concentrations is required for each cell line to identify optimal conditions where only a single spheroid is formed that is robust enough to allow manipulation without fragmenting. Optimally, the spheroids were between 200 to 500 µm in diameter, and consisted of tightly adherent cells with minimal cell debris. Spheroids survived gentle handling without damage, allowing them to be collected and used in a wide variety of assays.
Spheroid formation could be compromised by bacterial or other microbial contaminants (Figure 3A), which resulted in aggregates of dead cells. In the presence of dust or other fiber contamination, multiple or irregularly shaped cell clusters that were only weakly aggregated formed, and the resultant spheroids easily broke apart when handled. Spheroid formation was also affected by suboptimal concentrations of MC or serum in the spheroid formation medium (Figure 3B). In our testing, many cell lines were able to adhere to cell-repellent plates in the absence, or at low concentrations, of MC, and resulted in the formation of a spheroid surrounded by a cell monolayer (Figure 3B-ii). BxPC-3 cell growth in suboptimal MC conditions is shown as an example. In general, higher concentrations of MC prevented cells from adhering to the well, but too high a concentration of MC reduced cell-cell adhesion, and prevented cells from settling to the bottom of the well, resulting in the formation of loose aggregates and numerous satellite spheroids (Figure 3B-i). The concentration of serum also affected cell survival, and cell-cell, and cell-plastic adhesion and needed to be optimized for different cell lines. For cell lines such as HCT-116 and PANC-1, too high a serum concentration resulted in excessive cell proliferation and production of oversized spheroids that were easily damaged by handling, or promoted cell adhesion to the plastic well and the formation of a monolayer (Figure 3B-iii). Interestingly, the effects of insufficient serum differed between cell lines. HCT-116 cell survival was reduced and the spheroids formed were small, containing a large proportion of dead cells in low serum. In contrast, PANC-1 cells were viable in the absence of serum, but became more adherent, and formed multiple aggregates as well as a cell monolayer (Figure 3B-iv, v).
In invasion assays, pre-formed spheroids were resuspended in neutralized collagen to generate a rigid extracellular matrix (Section 2.2, Figure 4). Subsequent addition of a chemoattractant induced individual cells to move outwards from the spheroid and invade into the surrounding matrix (Figure 4B). The number of invading cells and distance invaded can be quantified through brightfield microscopy, or by fluorescence microscopy in the presence of a live cell stain such as DAPI or Calcein AM (Sections 3, 4). In our testing, morphological changes, such as the formation of protrusions, were visible within 6 h of treatment with chemoattractant, and numerous cells could be observed completely detaching from the spheroid and invading into the collagen within 18 to 48 h. For our experiments, we found that 12 to 18 h of invasion was ideal, as longer incubation times resulted in cells moving too far from the spheroid for optimal imaging. Fixed collagen-embedded spheroids could be imaged by brightfield, to quantify the distance and number of cells invading, or immunofluorescence microscopy (Sections 3-5), for visualization of invasive structures formed by the cells (Figures 5B, 5C).
A modification of the described spheroid formation protocol, in which individual cells are suspended in higher concentration MC (30 mg/mL) can be used to monitor anoikis resistance. At these MC concentrations, medium can still be transferred by pipette while cool, but thickened into a solid gel at 37 °C. Anoikis-resistant cells resuspended into this medium remained in suspension and proliferated over a period of 2-4 weeks, forming suspended spheroids of varying sizes (Figure 6B). Anchorage-dependent cells do not form spheroids under these conditions. We were able to quantify anoikis-resistance by counting the number of spheroids formed in each well.
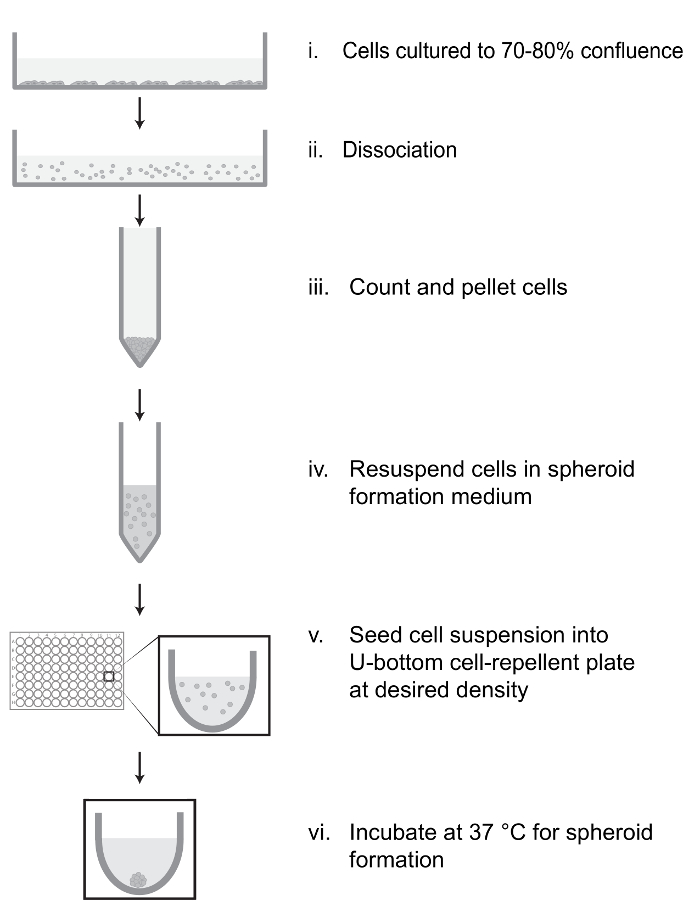
Figure 1: Overview of 3D Spheroid Formation Protocol. Cells grown in monolayer culture are dissociated, counted, pelleted, and resuspended in spheroid formation medium supplemented with methyl cellulose (MC) and serum (i-iv). The suspension is seeded into a U-bottom cell-repellent plate at desired density to allow formation of spheroids of the desired size (v), and the plate is incubated to produce a single, discrete cell spheroid in each well (vi). Please click here to view a larger version of this figure.
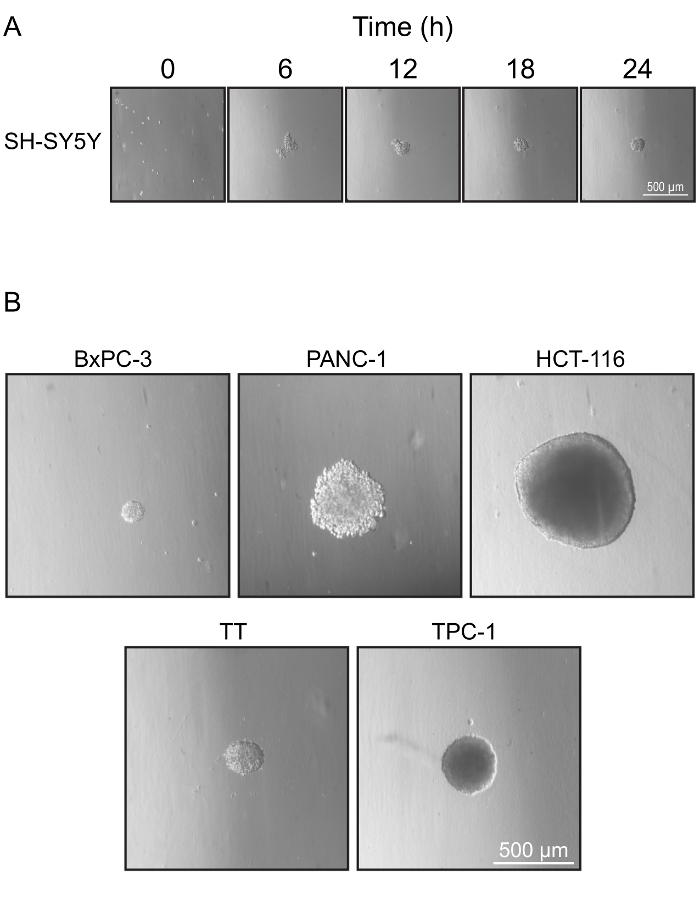
Figure 2: Optimized Spheroid Formation Conditions. (A) Spheroid formation under optimal conditions for SH-SY5Y cells. 1,000 cells seeded into U-bottom cell-repellent plates in media supplemented with 5% serum and 1 mg/mL MC aggregated into compact spheroids within 24 h. (B) Representative images of spheroids formed by various cell lines under optimal conditions shown in Table 1. Please click here to view a larger version of this figure.
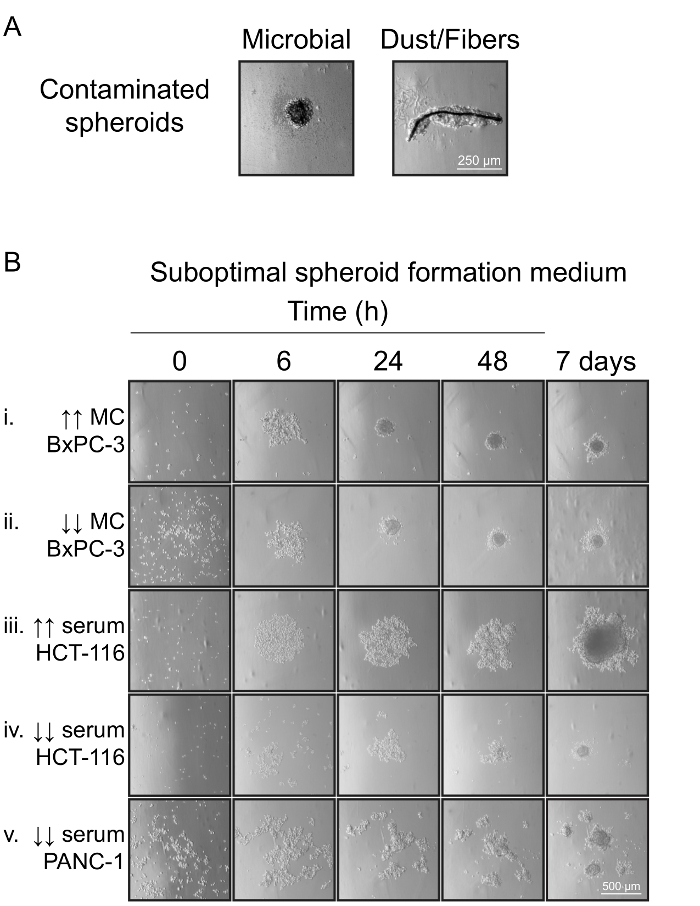
Figure 3: Spheroid Formation Under Suboptimal Conditions. (A) Spheroids formed in the presence of contaminants. Microbial contamination resulted in cell death and loosely aggregated spheroids. In the presence of insoluble contaminants, such as dust or fibers, cells adhered to these materials and did not aggregate. (B) Spheroid formation in suboptimal spheroid formation medium. BxPC-3 cells seeded in the presence of excess MC (10 mg/mL) formed numerous satellite spheroids (i), while insufficient MC (0 mg/mL) resulted in cell aggregates that adhered to the well bottom and formed a monolayer (ii). Similar results were observed for other tested cell lines (not shown). Excess serum (20%) resulted in increased HCT-116 cell proliferation and monolayer formation (iii). Insufficient serum (0%) had cell line-specific effects ranging from cell death in HCT-116 (iv) to the formation of satellite spheroids in PANC-1 (v). Please click here to view a larger version of this figure.
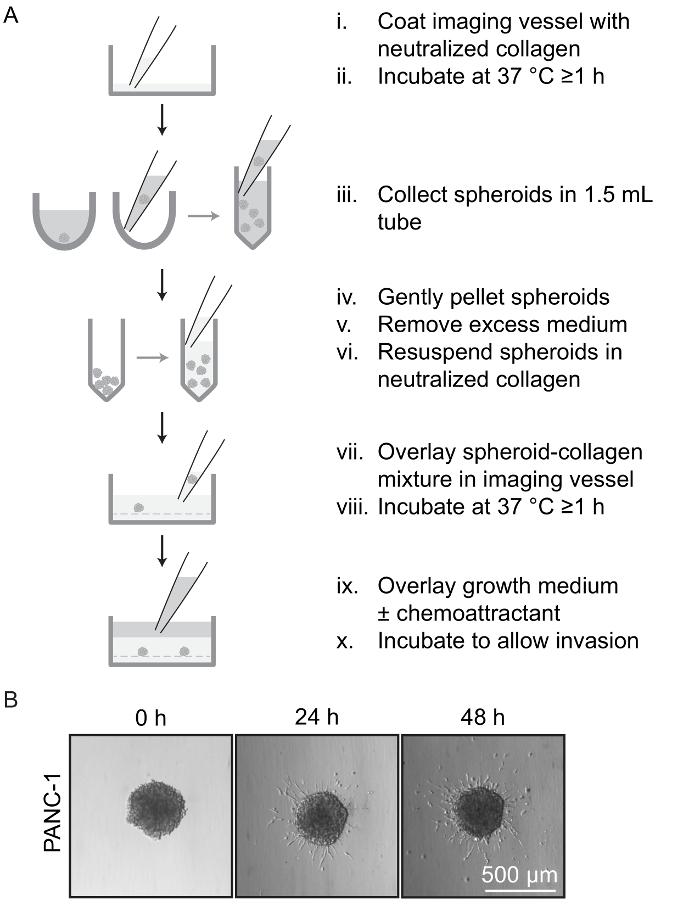
Figure 4: Invasion Assay Using Spheroids. (A) Overview of spheroid invasion assay. The imaging vessel is pre-coated with a layer of neutralized collagen and incubated to polymerize the collagen (i-ii). Pre-formed spheroids are collected in 1.5 mL tubes and resuspended in neutralized collagen (iii-vi). The spheroid-collagen mixture is overlaid on the collagen base-layer and incubated to polymerize the collagen (vii-viii). Growth medium containing desired compounds is then overlaid onto the polymerized spheroid-collagen layer and incubated to allow cell invasion (ix-x). (B) In the presence of a chemoattractant, cells are shown invading into the surrounding collagen matrix from an embedded PANC-1 spheroid at the indicated time points. Each panel shows a phase contrast image acquired using a 10X objective. Please click here to view a larger version of this figure.
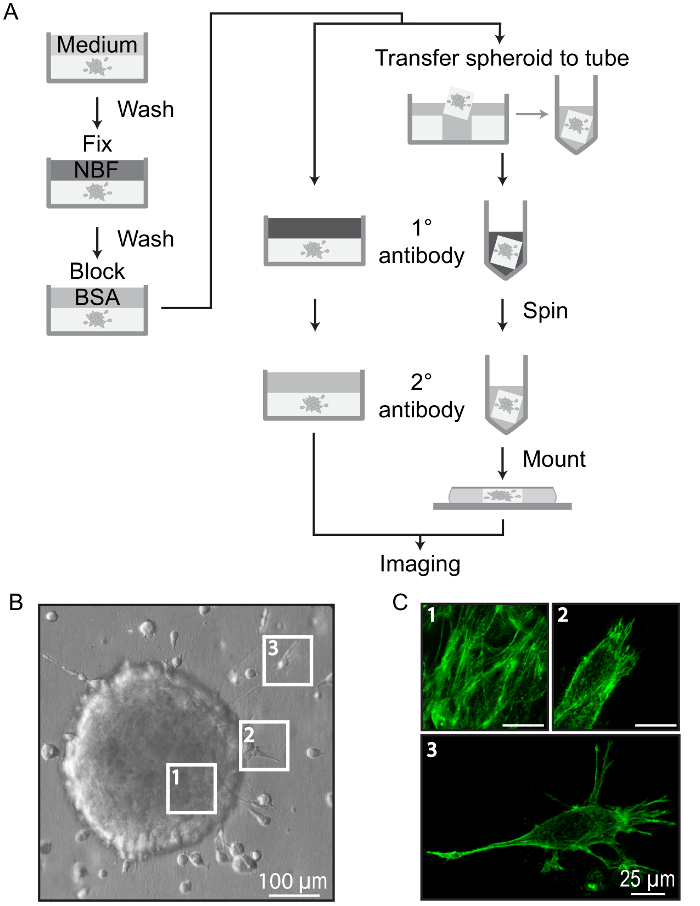
Figure 5: Immunofluorescence Staining of Collagen-embedded Spheroids. (A) Overview of protocol for immunofluorescence staining of spheroids. Embedded spheroids are washed, fixed (NBF), and blocked (BSA) directly in the imaging vessel. Spheroids can be stained directly, or excised and stained in a smaller volume, such as a 1.5 mL tube. Spheroids should be washed with excess buffer following each stain to minimize background fluorescence. Stained spheroids can be imaged directly in the vessel or mounted under a coverslip for imaging and storage. (B) Immunofluorescence images of a TPC-1 cell spheroid embedded in collagen and allowed to invade for 24 h. (C) Images showing phalloidin stained cells typical of regions indicated in B. Each panel shows a 20 µm Z-projection (0.2 µm steps) acquired using a 60X objective: Cells within the spheroid body approximately 20 µm from the surface (1), cells protruding into collagen (2), and cells invading through collagen (3). Scale bars = 25 µm. Please click here to view a larger version of this figure.
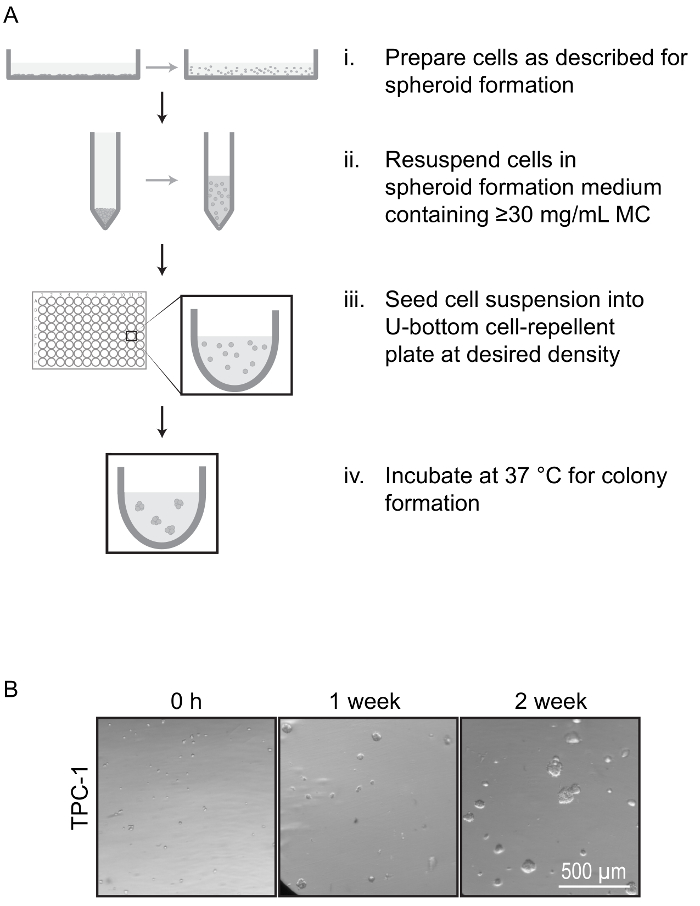
Figure 6: Spheroid Formation Anoikis Assay. (A) Overview of protocol for anoikis assay. Cells are prepared as described in Figure 1, but are resuspended in spheroid formation medium containing a minimum of 30 mg/mL MC which forms a thick layer to hold individual cells in suspension and prevent cell aggregation (i-ii). The cell suspension is seeded into U-bottom cell-repellent plates and incubated (iii-iv). The suspended cells can be monitored for spheroid formation by proliferation, indicating anoikis resistance. (B) Representative images of colony formation during anoikis assay. TPC-1 cells were held in suspension with 30 mg/mL MC and imaged at the indicated time points. Cells that are able to resist anoikis proliferate to form spheroids. Please click here to view a larger version of this figure.
| Cell Line | MC (mg/mL) | Serum (%) | Approximate incubation time |
| SH-SY5Y | 1 | 5 | 24 hours |
| BxPC-3 | 5 | 5 | 3 to 5 days |
| PANC-1 | 5 | 5 | 5 to 7 days |
| HCT-116 | 3 | 10 | 2 to 4 days |
| TT | 1 | 10 | 7 to 10 days |
| TPC-1 | 3 | 5 | 1 to 2 days |
Table 1: Optimal Spheroid Formation Medium Composition and Incubation Times for Validated Cell Lines.
