यह अनुमान है कि स्तनधारी जीनोम लगभग 20,000 प्रोटीन कोडिंग जीन होते हैं। वैकल्पिक स्प्लिसिंग और posttranslational संशोधनों आगे प्रोटीन प्रदर्शनों की सूची में वृद्धि। प्रोटीन एक मॉड्यूलर संरचना 1 है और अक्सर कई अन्योन्य क्रिया डोमेन है, जो विभिन्न प्रोटीन परिसरों में उनकी भर्ती और कई कोशिकीय प्रक्रियाओं 2 में उनकी भागीदारी के लिए अनुमति देने के होते हैं। एक उदाहरण बहुआयामी प्रोटीन p120ctn कहा जाता है। p120ctn Ctnnd1 जीन द्वारा इनकोडिंग और एक बड़े केंद्रीय वर्मी दोहराने डोमेन एक एन टर्मिनल और एक सी टर्मिनल क्षेत्र से घिरे होते हैं है। p120ctn की वर्मी डोमेन शास्त्रीय cadherins, जो सेल कोशिका आसंजन में शामिल हैं की एक अत्यधिक संरक्षित juxtamembrane डोमेन को बांधता है, लेकिन यह भी ट्रांस्क्रिप्शनल repressor Kaiso को बांधता है। p120ctn के एन टर्मिनल डोमेन अलग kinases, फास्फेटेजों, छोटे RhoGTPases, और सूक्ष्मनलिका जुड़े पी के साथ सूचना का आदान प्रदानroteins 3। दिलचस्प बात यह है विकल्प स्प्लिसिंग का एक परिणाम के रूप में, p120ctn isoforms चार विकल्प शुरू कोडोन 4 से उत्पन्न किया जा सकता है। के रूप में यह सबसे -5 से अनुवाद किया है कोडोन शुरू 'और पूर्ण लंबाई एन टर्मिनल खंड शामिल p120ctn isoform 1 ए, सबसे लंबे समय तक है। p120ctn में isoforms 3 और 4, इस एन टर्मिनल खंड आंशिक रूप से और पूरी तरह से क्रमश: हटाया जाता है। प्रोटीन (या प्रोटीन isoforms) और उनके डोमेन अलग सेलुलर कार्यों में की सटीक भूमिका को समझना एक चुनौती बनी हुई है।
जीन mESCs में लक्षित इसी जीन की आनुवंशिक विलोपन के माध्यम से एक प्रोटीन की कार्यप्रणाली के अध्ययन के लिए सक्षम बनाता है और व्यापक रूप developmentally महत्वपूर्ण और रोग-प्रासंगिक जीन और रास्ते की पहचान करने के लिए योगदान दिया है। रिवर्स आनुवंशिकी में यह सफलता समरूप पुनर्संयोजन 5 की वजह से mESC अलगाव और जीन लक्ष्यीकरण के क्षेत्र में प्रगति का परिणाम था </s> ऊपर। समरूप पुनर्संयोजन एक प्रक्रिया है जिसमें डीएनए टुकड़े दोहरे धागे (डी एस) डीएनए टूट जाता है के बाद दो या समरूप न्यूक्लिक moieties के बीच आदान-प्रदान किया जाता है। क्योंकि dsDNA टूटता निराला हैं आम तौर पर, मानव संसाधन अक्षम है। हाल ही में, अनुरूपता निर्देशित जीन लक्ष्यीकरण की दक्षता साइट विशिष्ट न्युक्लिअसिज़ 6, 7 का उपयोग कर बढ़ाया जा सकता है, लेकिन दुर्भाग्य से, इन ऑफ-टारगेट प्रभाव 8 से ग्रस्त हैं। एक और अधिक विश्वसनीय तकनीक जीन लक्ष्यीकरण सक्षम करने के लिए RMCE है, जो साइट विशिष्ट ऐसे Cre / loxP या FLPe / Frt के रूप में पुनर्संयोजन सिस्टम पर आधारित है। LoxP और Frt अनुक्रम जीवाणुभोजी P1 में पाए जाते हैं और Saccharomyces cerevisiae, क्रमशः, और 34 बीपी की, एक असममित 8 बीपी अनुक्रम है कि साइट के उन्मुखीकरण का निर्धारण करता है सहित मिलकर बनता है। दूसरी ओर,, के उन्मुखीकरण उदाहरण के लिए, एक डीएनए खिंचाव के भीतर दो loxP साइटों का निर्धारण करेगा floxed डीएनए excised हो जाता है कि क्या है या मैंCre की मध्यस्थता पुनर्संयोजन 9 पर nversed। इसके अलावा, यह भी एक रचनात्मक अनुवादन पैदा कर सकते हैं, तो दो साइटों अलग गुणसूत्रों पर स्थित हैं। RMCE heterospecific पुनर्संयोजन साइटों को नहीं पार प्रतिक्रिया करते हैं और है कि एक जीनोमिक ठिकाना में एम्बेडेड रहे हैं का लाभ लेता है। एक दाता प्लाज्मिड कि एक ही heterospecific साइटों से घिरे एक डीएनए टुकड़ा होता है की उपस्थिति में, recombinase डबल एक साथ अनुवादन (चित्रा 1) की वजह से RMCE संगत जीनोमिक ठिकाना में इस डीएनए टुकड़ा डाल देंगे। इधर, केवल सही ढंग से RMCE-लक्षित क्लोन भेजे वेक्टर पर एक प्रमोटर कि पुनर्स्थापित करने के लिए दवा प्रतिरोध धन्यवाद प्रदान कर सकते हैं एक "फंस" प्रमोटर कम Neomycin प्रतिरोध जीन (नव आर) डॉकिंग कोशिकाओं के R26 जीनोम में मौजूद (चित्रा 1) 10, 11। यह एक बहुत ही उच्च लक्ष्य-निर्धारण दक्षता, अक्सर 100% 11 के करीब है, में जो परिणाम </ sup> 12। अंत में, RMCE-आधारित लक्ष्य अत्यधिक कुशल है और संरचना-कार्यों के अध्ययन के लिए इस्तेमाल किया जा सकता; तथापि, यह एक पूर्व इंजीनियर जीनोमिक ठिकाना की आवश्यकता है।
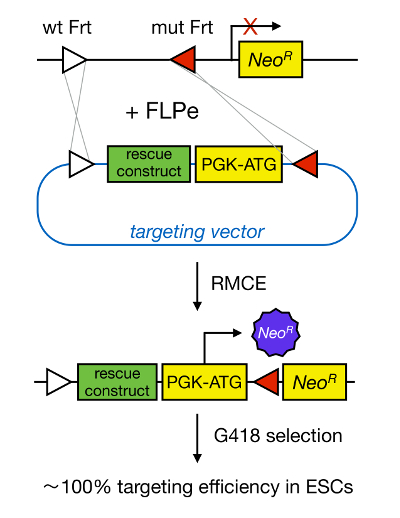
चित्र 1 RMCE की मध्यस्थता लक्ष्य निर्धारण के योजनाबद्ध प्रतिनिधित्व। अगर दोनों दो heterospecific Frt साइटों (सफेद और लाल त्रिकोण द्वारा दर्शाया) बंदरगाह RMCE एक परिभाषित जीनोमिक ठिकाना को भेजे को लक्षित वेक्टर से डीएनए क्षेत्रों के आदान प्रदान के लिए अनुमति देता है। इसके अलावा, इंजीनियर जीनोमिक ठिकाना एक promoterless और छोटा कर दिया neomycin प्रतिरोध (नव आर) जीन होता है। भेजे डीएनए टुकड़ा में एक प्रमोटर कोडोन प्रदान करने और शुरू से, केवल सही पुनर्संयोजन घटनाओं neomycin प्रतिरोध बहाल, उच्च लक्ष्य-निर्धारण क्षमता में जिसके परिणामस्वरूप। टी का एक बड़ा संस्करण देखने के लिए यहां क्लिक करेंउसके आंकड़ा।
mESCs में जीनोम इंजीनियरिंग RMCE संगत चूहों की पीढ़ी के लिए अनुमति देता है। 1981 में, दो समूहों ब्लास्टोसिस्ट की आंतरिक कोशिका द्रव्यमान (आईसीएम) से pluripotent कोशिकाओं पर कब्जा करने में और उन्हें संस्कृति 13, 14 में बनाए रखने में सफल रहा। mESCs भ्रूण और वयस्क कोशिकाओं के सभी प्रकार, जर्म सेल वंश सहित में आत्म नवीकरण और भेदभाव करने में सक्षम हैं। इसलिए, जीन mESCs में लक्षित संघटित या सशर्त (Cre / LoxP प्रणाली का उपयोग कर) को चूहों के विकास के माध्यम से रिवर्स आनुवंशिक अध्ययन सक्षम बनाता है। हालांकि, शास्त्रीय तरीका माउस ES कोशिकाओं को अलग करने के बहुत अक्षम है। कई प्रमुख सुधार बहुत पाने mESC लाइनों के लिए सफलता की दर में वृद्धि हुई है, एक परिभाषित सीरम बदलने (एसआर) मध्यम 15 के उपयोग सहित, mESC मध्यम के बीच अदल-एसआर और भ्रूण गोजातीय सीरम (एफबीएस) 16 युक्त, और फार्माको के उपयोगइस तरह के pluripotin या 2i 17 के रूप में तार्किक यौगिकों। Pluripotin, एक छोटे से सिंथेटिक अणु, ल्यूकेमिया निरोधात्मक कारक (LIF) और माउस भ्रूण fibroblasts (MEFs) 18 के अभाव में एक undifferentiated राज्य में mESCs के प्रसार के लिए अनुमति देता है। अंत में, यह दिखाया गया है कि mESCs एक बहुत ही उच्च दक्षता (100% के करीब) एक एसआर / एफबीएस मध्यम प्रत्यावर्तन प्रोटोकॉल, LIF के साथ और 19 pluripotin 20 संयुक्त है जब साथ अलग किया जा सकता। इन प्रोटोकॉल RMCE-संगत को mESCs कि बाद में संरचना-समारोह के अध्ययन के लिए इस्तेमाल किया जा सकता की कुशल अलगाव सक्षम करें।
इस पत्र के लिए एक विधि है कि एक एक प्रोटीन है कि विशिष्ट कोशिकीय प्रक्रियाओं के लिए जिम्मेदार हैं के भीतर प्रमुख डोमेन या अवशेषों की पहचान करने के लिए सक्षम बनाता है वर्णन करता है। इस उद्देश्य के लिए उन्नत प्रौद्योगिकियों कि कुशल mESC अलगाव सक्षम, को लक्षित वेक्टर विधानसभा, और mESC लक्ष्य-निर्धारण की एक पाइप लाइन बनाने थाघ। प्रोटीन isoforms, डोमेन उत्परिवर्ती, और नीचे की ओर प्रभावोत्पादक के साथ इस तरह, बड़े पैनल को mESCs में पेश किया जा सकता है और इन विट्रो KO phenotype बचाव करने की क्षमता के लिए मूल्यांकन किया जा सकता है।
