Se estima que los genomas de mamíferos contienen alrededor de 20.000 genes codificadores de proteínas. Splicing alternativo y modificaciones postraduccionales aumentan aún más el repertorio de proteínas. Las proteínas tienen una estructura modular 1 y a menudo contienen varios dominios de interacción, que permiten su reclutamiento en diferentes complejos de proteínas y su participación en múltiples procesos celulares 2. Un ejemplo es la proteína multifuncional llamada p120ctn. p120ctn está codificada por el gen Ctnnd1 y consta de un gran dominio central armadillo repetición flanqueado por un N-terminal y una región C-terminal. El dominio armadillo de p120ctn se une a un dominio yuxtamembrana altamente conservada de cadherinas clásicas, que están implicadas en la adhesión célula-célula, pero también se une al represor transcripcional Kaiso. El dominio N-terminal de p120ctn interactúa con diferentes quinasas, fosfatasas, pequeñas RhoGTPases, y asociada a los microtúbulos proteins 3. Curiosamente, como resultado de corte y empalme alternativo, isoformas p120ctn pueden ser generados a partir de cuatro codones de inicio alternativos 4. p120ctn isoforma 1A es la más larga, ya que se traduce de la más-5' codón de inicio y contiene el segmento N-terminal de longitud completa. En p120ctn isoformas 3 y 4, este segmento N-terminal se suprime parcialmente y completamente, respectivamente. La comprensión del papel exacto de proteínas (o isoformas de proteínas) y sus dominios en diferentes funciones celulares sigue siendo un reto.
La orientación de genes en mESCs permite el estudio de la función de una proteína a través de la supresión genética del gen correspondiente y ha contribuido ampliamente a la identificación de genes y las vías de desarrollo importantes y relevantes de la enfermedad. Este avance en la genética inversa fue el resultado de los avances en los campos de aislamiento mESC y la orientación de genes debido a la recombinación homóloga 5 </sarriba>. La recombinación homóloga es un proceso en el que fragmentos de ADN se intercambian entre dos restos nucleicos similares o idénticos después de doble hebra (ds) se rompe de ADN. Normalmente, HR es ineficiente debido a roturas de ADN bicatenario son poco frecuentes. Recientemente, la eficiencia del gen homología dirigida de orientación podría aumentarse usando nucleasas específicas de sitio 6, 7, pero, por desgracia, estos son propensos a los efectos fuera del objetivo 8. Una técnica más fiable para permitir la orientación de genes es RMCE, que se basa en los sistemas de recombinación específica de sitio tales como Cre / loxP o FLPe / FRT. LoxP y la secuencia Frt se encuentran en bacteriófago P1 y Saccharomyces cerevisiae, respectivamente, y se componen de 34 pares de bases, incluyendo una secuencia de pares de bases asimétrica 8 que determina la orientación del sitio. Por otra parte, la orientación de, por ejemplo, dos sitios loxP dentro de un tramo de ADN determinará si el ADN floxed convierte escindió o inversed tras la recombinación mediada por Cre 9. Por otra parte, Cre también puede inducir una translocación si dos sitios están localizados en diferentes cromosomas. RMCE se aprovecha de sitios de recombinación heterospecific que no reaccionan de forma cruzada y que están incrustados en un locus genómico. En presencia de un plásmido donante que contiene un fragmento de ADN flanqueado por los mismos sitios heterospecific, la recombinasa se insertar este fragmento de ADN en el locus genómico compatible-RMCE debido a la translocación de doble simultáneo (Figura 1). Aquí, los clones solamente correctamente orientados RMCE pueden hacer de resistencia a fármacos gracias a un promotor en el vector entrante que restaura una "atrapado" promotor-menos gen de resistencia a neomicina (Neo R) presente en el genoma R26 de las células de conexión (Figura 1) 10, 11. Esto resulta en una eficiencia muy alta focalización, a menudo cerca de 100% 11, </ sup> 12. En conclusión, la selección de base RMCE es altamente eficiente y se puede utilizar para estudios de estructura-funciones; sin embargo, se requiere un locus genómico pre-ingeniería.
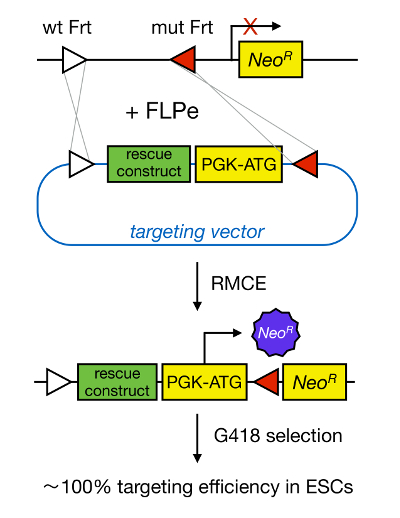
Figura 1. Representación esquemática de la orientación de RMCE mediada. RMCE permite el intercambio de segmentos de ADN a partir de un vector de dirección entrante a un locus genómico definido si ambos albergan dos sitios FRT heterospecific (representados por triángulos blanco y rojo). Además, el locus genómico de ingeniería contiene un gen truncado de resistencia a neomicina (Neo R) sin promotor y. Al proporcionar un promotor y el codón de inicio en el fragmento de ADN entrante, eventos de recombinación solamente correctas restaurar la resistencia a neomicina, dando como resultado altas eficiencias de focalización. Haga clic aquí para ver una versión más grande de tsu figura.
la ingeniería del genoma en mESCs permite la generación de ratones compatible-RMCE. En 1981, dos grupos tuvieron éxito en la captura de células pluripotentes de la masa celular interna (ICM) de blastocistos y en el mantenimiento de ellos en la cultura 13, 14. mESCs son capaces de auto-renovación y diferenciación en todos los tipos de células embrionarias y adultas, incluyendo el linaje de células germinales. Por lo tanto, la orientación de genes en mESCs permite estudios inversa genéticos mediante el desarrollo de ratones KO constitutivos o condicionales (utilizando el sistema cre / loxP). Sin embargo, la manera clásica para aislar células ES de ratón es muy ineficiente. Varias mejoras importantes han aumentado en gran medida la tasa de éxito para derivar líneas mESC, incluyendo el uso de un suero de reemplazo definido (SR) medio 15, alternando entre medio mESC que contenía suero bovino SR y fetal (FBS) 16, y el uso de fármacocompuestos lógicos como pluripotin o 2i 17. Pluripotin, una molécula sintética pequeña, permite la propagación de mESCs en un estado no diferenciado en ausencia del factor inhibidor de la leucemia (LIF) y fibroblastos embrionarios de ratón (MEFs) 18. Por último, se ha demostrado que mESCs se pueden aislar con una eficacia muy alta (cerca de 100%) cuando un / FBS protocolo alternancia medio SR se combina con LIF y pluripotin 19, 20. Estos protocolos permiten el aislamiento eficiente de mESCs KO compatible con RMCE que posteriormente pueden ser utilizados para estudios de estructura-función.
Este documento describe un método que permite identificar los dominios o residuos clave dentro de una proteína que son responsables de los procesos celulares específicos. Con este fin, una línea de tecnologías avanzadas que permiten el aislamiento eficiente mESC, montaje vector de dirección y orientación mESC era crearre. Como tales paneles grandes, con isoformas de proteínas, mutantes de dominio, y efectores aguas abajo pueden ser introducidas en mESCs KO y se pueden evaluar por su capacidad para rescatar el fenotipo vitro KO en.
