Mammary VESCs Isolation:
To isolate the endothelial population for the consequent transplantation assay, mature (8 week-old), virgin mouse mammary glands were harvested as donor material. Endothelial cells were isolated using an antibody-based cell sorting technique. Representative plots of the FACS analysis of the VESC population in 8-week-old C57BL/6 mammary gland ECs are shown in Figure 1 (Figure adapted from Yu et al.8, with permission).
Mammary Fat Pad Transplantation:
The mammary fat pad provides an ideal site for the study of angiogenesis. After FACS-based isolation, Procr+ ECs were mixed with 0.5% basement membrane matrix and 0.1% Trypan blue indicator and were then transplanted to the empty fat pad of 3-week-old SCID recipients. During puberty, the mammary fat pad creates an angiogenic environment, promoting the remodeling of the vasculature in alignment with mammary epithelium expansion. As a result, transplanted Procr+ ECs (GFP+) incorporated into the large vessels of the host (Figure 2B, arrows) and also formed secondary and tertiary GFP+ vessel branches (Figure 2B, arrowheads). Although transplanted Procr+ ECs can also generate vessels in a matrix plug assay (inserted under the flank skin), the formed vessels only appear capillary-like (Figure 2A; figure adapted from Yu et al.8, with permission).
Functionality of the Vessels Formed by Procr+ VESCs:
To investigate whether the GFP+ vessels in mammary fat pads are part of the functional circulatory vasculature, isolectin was administered by intravenous injection prior to harvest. The GFP+ vessels were isolectin+, indicating that the formed vessels are luminized and connected with the host vasculature (Figure 2B; figure adapted from Yu et al.8, with permission).
Mammary VESC Lineage Tracing Using the ProcrCreERT2; Rosa26mTmGMouse Model:
ProcrCreERT2; Rosa26mTmG mice were used for the fate tracing of Procr+ VESCs in vivo (Figure 3A). To investigate how Procr+ VESCs contribute to the robust angiogenesis and vascular remodeling during mammary gland pubertal development, ProcrCreERT2; Rosa26mTmG were administered with tamoxifen to induce GFP expression in the Procr+ endothelial population. The mammary fat pads of drug-treated ProcrCreERT2; Rosa26mTmG mice were harvested both short-term (2 days post-injection; Figure 3B) and long-term (up to 10 months post-injection; Figure 3C-F; figure adapted from Yu et al.8, with permission). The whole-mount preparation was carried out to visualize the tracing outcome. The vascular endothelium identity can be validated by staining with endothelial surface marker VE-Cadherin (Cdh5; Figure 3B, right image).
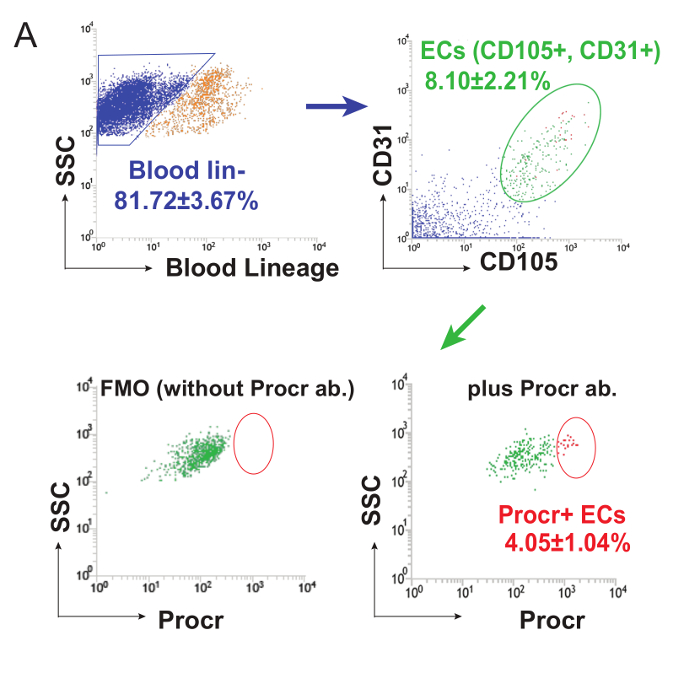
Figure 1: Analysis of VESCs in Mouse Mammary Gland. FACS analysis of Procr expression in 8-week-old C57BL/6 mammary gland ECs. 8 week-old virgin C67BL/6 female mice were used for mammary gland collection. After cell samples were prepared, unstained sample and each single-color control were analyzed to determine the voltage for each color. The compensation parameters were calculated to avoid auto-fluorescence or background staining. A template was established for proper gating and to sort for the desired cell populations. To establish the hierarchy of FACS sorting, debris from all events was eliminated and then doublets or adhesive cell clusters were discarded. Lineage- cells were gated out, and CD31 and CD105 were used to define the endothelial populations. Procr+ cells were isolated from CD31+ CD105+ ECs compared to FMO control. This figure has been modified from Yu et al.8, with permission. Please click here to view a larger version of this figure.
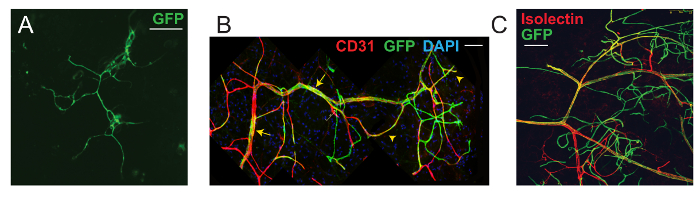
Figure 2: VESC Mammary Fat Pad Transplantation Assay. (A) A vessel formed from Procr+ ECs within a basement membrane matrix plug inserted subcutaneously under the flank skin. (B) Confocal image of a recipient mammary fat pad, indicating the integration and contribution of transplanted Procr+ ECs (GFP+) to host mammary vasculature. ECs were counterstained with CD31. (C) Intravenous injection in recipient mice with isolectin showed that the outgrowths formed luminized vessels. Scale bar, 50 µm. Images were acquired using a 40X N.A. 1.3 oil objective. Tile scans of Z-stacks were acquired at an optical section resolution of 1,024 x 1,024, and each layer was 1.0 – 1.2 µm apart. To capture the correct fluorescence light signal, the filter beam splitter was set to TD 488/553/638, with the gain setting at PMT Gain 600 – 800. The following excitation/emission sources were used: mGFP, with excitation/emission maxima of 488/509 nm; mTomato, with excitation/emission maxima of 532/588 nm; Alexa-647, with excitation/emission maxima of 594/665; and DAPI, with excitation/emission maxima of 350/470. This figure has been modified from Yu et al.8, with permission. Please click here to view a larger version of this figure.
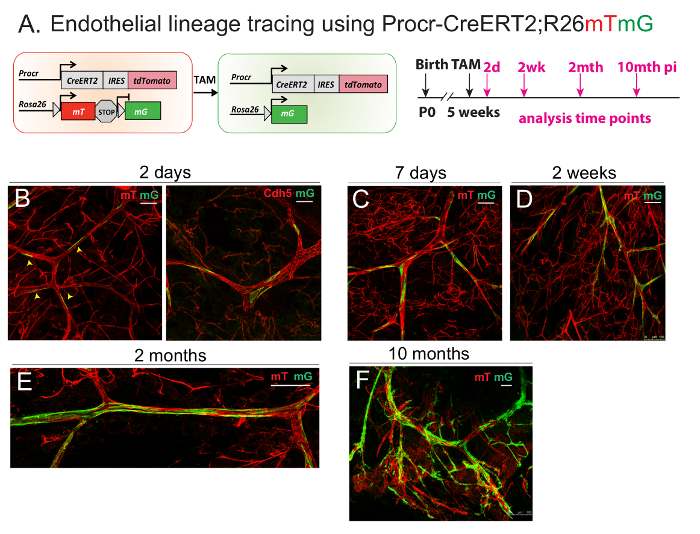
Figure 3: Lineage Tracing of Procr+ ECs Demonstrates their Contribution to EC Clonal Expansion During Development. (A) Illustration of the lineage tracing strategy using ProcrCreERT2; Rosa26mTmG line (Left panel). Experimental setup used in the short-term (2 days) and long-term (7 days to 10 months), as indicated (Right panel). (B-F) Whole-mount confocal imaging of mammary vasculature after different lengths of tracing periods, indicating the initial location of labeled Procr+ ECs and their contribution towards mammary vasculature at various tracing stages. Scale bar = 50 µm. This figure has been modified from Yu et al.8, with permission. Please click here to view a larger version of this figure.
