केंद्रीय तंत्रिका तंत्र (सीएनएस) एक सीमित क्षमता को नुकसान और/या न्यूरॉन्स और axonal मार्ग है कि ऐसे दर्दनाक मस्तिष्क की चोट (TBI), स्ट्रोक, रीढ़ की हड्डी की चोट (विज्ञान), और neurodegenerative रोग के रूप में शर्तों के साथ की शिथिलता प्रभावहीन है1 ,2,3,4,5. सीएनएस में Neurogenesis मस्तिष्क में क्षेत्रों की एक सीमित संख्या के लिए प्रतिबंधित है, खो ंयूरॉंस की बहाली में बाधा6,7। इसके अतिरिक्त, सीएनएस में खो axonal रास्ते का पुनर्जनन निर्देशित मार्गदर्शन की कमी के कारण अपर्याप्त है, वृद्धि अवरोधकों की उपस्थिति, और प्रतिक्रियाशील astrogliosis तंत्रिका ऊतक के लिए निंनलिखित क्षति2,8, 9,10. Astrocytes आमतौर पर आयन homeostasis, न्यूरोट्रांसमीटर मंजूरी, synapse गठन, और neurovascular युग्मन के साथ न्यूरॉन्स की सहायता करने में विविध कार्य किया है11. फिर भी, तंत्रिका ऊतक को हल्के नुकसान के बाद, astrocytes आणविक, संरचनात्मक, और कार्यात्मक परिवर्तन से गुजरना हो सकता है के रूप में वे एक hypertrophic राज्य के लिए संक्रमण11। गंभीर neurotrauma के जवाब में, इन परिवर्तनों के परिणामस्वरूप एक penumbra के साथ एक निशान के गठन में प्रतिक्रियाशील astrocytes और एक घाव कोर जिसमें टूटना रक्त-मस्तिष्क बाधा से लीक ल्यूकोसाइट्स शामिल है (BBB), microglia, oligodendrocytes, व fibroblasts११,१२,१३. ये प्रतिक्रियाशील astrocytes, मध्यम रेशा प्रोटीन और chondroitin सल्फेट proteoglycans (CSPGs), जो तंत्रिका पुनर्जनन12में बाधा की बढ़ती अभिव्यक्ति की एक आकृति विज्ञान, अव्यवस्थित प्रक्रियाओं और प्रदर्शनी की वृद्धि हुई । हालांकि glial निशान शुरू में मदद करता है BBB अखंडता को बहाल करने और आसपास के स्वस्थ ऊतकों के लिए भड़काऊ प्रतिक्रिया के संचरण से बचने, यह axon पुनर्जनन12के खिलाफ एक भौतिक और जैव रासायनिक बाधा के रूप में कार्य करता है,14 ,15,16. उदाहरण के लिए, axons है कि मुठभेड़ glial निशान प्रदर्शन बल्ब अपक्षयी विकास शंकु और स्टंट विकास12। इसके अलावा, astrocytic की प्रक्रिया की चोट के बाद axons17पुनः उत्पन्न करने के विस्तार में बाधा उत्पन्न करता है । इन निरोधात्मक विशेषताओं का परिणाम अक्सर स्थाई शारीरिक और स्नायविक दोष है कि रोगियों को गंभीर neurotrauma, TBI और विज्ञान सहित के बाद पीड़ित में प्रकट होता है ।
बाह्य सीएनएस में कार्यात्मक उत्थान का सामना करना पड़ रहा चुनौतियों के बावजूद, axons को पुनर्जीवित करने के लिए एक आंतरिक क्षमता के अधिकारी के लिए दिखाया गया है । उदाहरण के लिए, glial निशान के साथ संपर्क में अपक्षयी विकास शंकु के गतिशील प्रकृति पता चलता है कि इन अंत अपनी क्षमता को बनाए रखने के लिए12का विस्तार । नतीजतन, यह माना जाता है कि axonal पुनः वृद्धि के लिए एक मुख्य बाधा पोस्ट सीएनएस के निरोधात्मक वातावरण है और यह है कि glial scarring को कम करने के माध्यम से एक अधिक स्वतंत्र वातावरण प्रदान करने और/या निशान भर में अपक्षयी पुलों प्रदान होगा लाभप्रद. दरअसल, पिछले अध्ययनों का प्रदर्शन किया है कि सीएनएस न्यूरॉन्स पुलों के रूप में परिधीय तंत्रिका भ्रष्टाचार का उपयोग कर एक घाव के माध्यम से axons का विस्तार करने में सक्षम थे, जो axon पुनर्जनन12के लिए एक और अधिक अनुकूल वातावरण वर्तमान,18, 19. कई अंय रणनीतियों इस vestigial अपक्षयी क्षमता का दोहन करने के लिए पीछा किया गया है । उदाहरण के लिए, सेल विकास के विभिंन चोट मॉडलों में संकेत रास्ते के हेरफेर axonal पुनर्जनन और glial निशान कमी10,20,21में हुई है । इसके अतिरिक्त, अध्ययनों से पता चला है कि chondroitinase एबीसी, जो CSPGs में चीनी श्रृंखला के बहुमत से सट के साथ उपचार, CSPGs के निरोधात्मक प्रभाव को कम करने के लिए सक्रिय द्वारा स्रावित astrocytes22। उत्साहजनक परिणाम के बावजूद, इन तरीकों विकास शंकु, जो संभवतः ंयायपालिका पुनर्जनन12में परिणाम कर सकते है के मार्गदर्शन निर्देशित प्रदान नहीं करते हैं, और भी ंयूरॉंस के नुकसान के लिए खाते में नहीं है । सेल आधारित दृष्टिकोण glial निशान के प्रभाव को दुर्गम और खो कोशिकाओं, विशेष रूप से ंयूरॉंस भरपाई करने के प्रयास में उपयोग किया गया है । कुछ समूहों astrocytes न्यूरॉन्स में विभेदित प्रतिक्रियाशील है, जबकि दूसरों को चोट क्षेत्र को फिर से बसाने और axon पुनर्जनन को बढ़ावा देने के लिए सीएनएस घावों में तंत्रिका जनक कोशिकाओं प्रत्यारोपण किया है23,24, 25. हालांकि, स्टेम सेल प्रत्यारोपण अकेले कम जीवित रहने की दर, गरीब एकीकरण, और क्षतिग्रस्त ऊतक में मामूली प्रतिधारण द्वारा सीमित है5. इसके अलावा, इन सेल आधारित रणनीतियों के लिए लंबी दूरी axonal इलाकों, विशेष रूप से एक नियंत्रित तरीके से बहाल करने में विफल । इसलिए, अन्य दृष्टिकोण के साथ संयोजन में, विभिन्न तंत्रिका और जनक कोशिकाओं और विकास कारकों के लिए वितरण वाहनों के रूप में पता लगाया जा रहा है26। डिजाइन नियंत्रण के एक उच्च डिग्री-आधारित दृष्टिकोण सुविधा का निर्माण करने के लिए कि विशिष्ट शारीरिक, haptotaxic, और chemotaxic तीन आयामी (3 डी) लक्ष्य मेजबान ऊतक के microenvironment में मौजूद cues नकल का उत्पादन करने के लिए27, 28,29,30,31,३२,३३,३४. इन पर्यावरणीय संकेतों के प्रजनन, प्रत्यारोपण कोशिकाओं के लिए सर्वोपरि है मूल आकृति विज्ञान, प्रसार, प्रवास, और संकेतन, अन्य neurobiological विशेषताओं के बीच29पेश करने के लिए । इन लाभप्रद गुणों के बावजूद, पारंपरिक कोशिका से परे उंनति के लिए बीज बोने की एक साथ लंबी दूरी की axonal पुनर्जनन निर्देश को बढ़ावा देने और खो ंयूरॉंस की जगह की आवश्यकता है ।
एक आशाजनक वैकल्पिक दृष्टिकोण पर आधारित है तंत्रिका ऊतक इंजीनियर “रहने पाड़”, जो अन्य कोशिका से अलग कर रहे हैं आधारित एक अनुरूप cytoarchitecture के साथ तंत्रिका कोशिकाओं के रहने की उपस्थिति के कारण दृष्टिकोण है कि देशी neuroanatomy का अनुकरण और/ विकासात्मक तंत्र लक्षित प्रतिस्थापन, पुनर्निर्माण, और तंत्रिका सर्किट4के उत्थान की सुविधा के लिए,३५। पाड़ों के डिजाइन के लिए विचार phenotypes और तंत्रिका कोशिकाओं के स्रोतों, साथ ही साथ यांत्रिक/भौतिक संपत्तियों और जैव रासायनिक संकेतों के किसी भी साथ के संयोजन से तय की संरचना में शामिल है३५। इन विट्रो मेंनिर्माण के बाद, इन रहने वाले पाड़ vivo में सेल-आसंजन अणुओं और chemotactic और neurotrophic संकेतों को सक्रिय रूप से विनियमित तंत्रिका सेल प्रवासन और राज्य के आधार पर axon वृद्धि को नियंत्रित करने के लिए प्रत्यारोपित किया जा सकता है और ३५reअपक्षय प्रक्रियाओं की प्रगति । Glial कोशिकाओं के रहने वाले पाड़ के इंजीनियर cytoarchitecture के लिए एक आधार के रूप में इन कोशिकाओं vivo मेंविभिंन विकासात्मक तंत्र मध्यस्थता के बाद सेवा कर सकते हैं । मस्तिष्क के विकास के दौरान, नए ंयूरॉंस बेसल निर्देशित प्रवासन३६,३७के लिए रहने वाले पाड़ के रूप में विकासशील cortical प्लेट की ओर वेंट्रिकुलर क्षेत्र से रेडियल glia द्वारा विस्तारित प्रक्रियाओं पर निर्भर हैं । इसके अलावा, विकास शंकु का विस्तार करने के लिए खुद को आकर्षक और क्रीमी glial guidepost कोशिकाओं द्वारा उठाए गए संकेतों संवेदन द्वारा उंमुख दिखाया जाता है, और तथाकथित “अग्रणी” axons को पूर्व पैटर्न के साथ विस्तार से सही लक्ष्य तक पहुंचने का सुझाव दिया जाता है glial पाड़ो३५,३८,३९. इस प्रकार, glial कोशिकाओं अग्रणी axons के मार्गदर्शन के लिए आवश्यक हैं, जो बाद में axon के रूप में सेवा आधारित “रहने पाड़” के प्रक्षेपण निर्देशित करने के लिए “अनुयाई” axons । इसके अलावा, glia-मध्यस्थता विकास तंत्र postnatally बनाए रखने के लिए दिखाया गया है, के रूप में neuroblasts rostral प्रवासी धारा (RMS) का पालन करने के लिए subventricular क्षेत्र से नेविगेट करने के लिए (SVZ), वयस्क मस्तिष्क में neurogenesis के कुछ शेष क्षेत्रों में से एक, घ्राण बल्ब (ओबी)४० RMS में इन neuroblasts glial ट्यूब (चित्र 1a-1), जो longitudinally-संरेखित astrocytic प्रक्रियाओं, सीधे सेल-सेल आसंजन और स्थानीयकृत घुलनशील कारकों३७के माध्यम से शामिल है के भीतर माइग्रेट करें, ४१. अंत में, जबकि स्तनधारियों में सीएनएस नुकसान बाधित astrocytic प्रक्रिया एक glial निशान है कि शारीरिक रूप से axonal पुनर्जनन17, कई गैर स्तनधारी प्रणालियों एक हानिकारक glial निशान के गठन की कमी के गठन की व्यवस्था का कारण बनता है । बल्कि, गैर स्तनधारी प्रजातियों के glial कोशिकाओं को और अधिक संगठित, गठबंधन पैटर्न है कि घायल क्षेत्र के माध्यम से गाइड के रूप में उपयोग किया जाता है बनाए रखने के17,४२,४३। उदाहरण के लिए, गैर में स्तनधारी विज्ञान मॉडल, axons घाव पार glial पुलों के साथ निकट सहयोग में विकसित करने के लिए दिखाया जाता है, axonal पुनर्जनन और कार्यात्मक वसूली को सुविधाजनक बनाने के रूप में आयोजित glial पाड़ों के लिए एक महत्वपूर्ण भूमिका का सुझाव ( चित्र 1a -2) ४२ , ४४ , ४५. neuroanatomical सुविधाओं के Recapitulation और विकासात्मक/ऊपर वर्णित इंजीनियर glial के एक नए वर्ग उपज हो सकता है आधारित रहने वाले पाड़ों कि समवर्ती ड्राइव कर सकते है आवर्तक ंयूरॉन प्रवासन और axonal pathfinding अंयथा गैर स्वतंत्र वातावरण के माध्यम से, जिससे संभावित सीएनएस चोट और रोग के साथ जुड़े और axon पथ अध…
हमारे अनुसंधान समूह पहले से पुनर्निर्माण और सीएनएस में axonal इलाकों के उत्थान और परिधीय तंत्रिका तंत्र (पीएन) के माध्यम से सूक्ष्म ऊतक इंजीनियर तंत्रिका नेटवर्क (माइक्रो-टेननस) और ऊतक के लिए रहने वाले पाड़ के कई प्रकार के डिजाइन किए है इंजीनियर तंत्रिका भ्रष्टाचारियों (TENGs), क्रमशः27,४६,४७,४८। दोनों रणनीतियों स्वाभाविक रूप से नकल पर आधारित हैं । माइक्रो-टेननस शारीरिक-प्रेरित संरचनाओं को संरचनात्मक रूप से डिज़ाइन किया गया है और कार्यात्मक रूप से मस्तिष्क के विशिष्ट न्यूरॉन आबादी को जोड़ने वाले axonal इलाकों को प्रतिस्थापित करते हैं । TENGs axon के विकासात्मक तंत्र का दोहन-axonal पुनर्जनन की सुविधा, उदाहरण “के साथ अनुयाई” axon विकास के साथ “पायनियर” axons, लक्षित मेजबान axonal पुनर्जनन प्राप्त करने के लिए३५,४६,४८। हम हाल ही में रहने पाड़ तकनीक की बहुमुखी प्रतिभा पर कैपिटल माइक्रो के रूप में एक समान झालर योजना का उपयोग-टेननस और glia से प्रेरणा की मांग आधारित विकास भर में मौजूद तंत्र । यहां, हम विकसित गठबंधन astrocytic एक hydrogel माइक्रो कॉलम४९के कोलेजन लुमेन फैले बंडलों से मिलकर निर्माण । इन astrocytic रहने वाले पाड़ पहले एक केशिका ट्यूब को भरने से विकसित कर रहे हैं-तरल agarose के साथ एक्यूपंक्चर सुई विधानसभा एक बाहरी व्यास (आयुध डिपो) और भीतरी व्यास (आईडी) के व्यास के लिए इसी के साथ एक खोखले बेलनाकार hydrogel बनाने के लिए ट्यूब और सुई, क्रमशः । agarose जमाना और hydrogel केशिका ट्यूब से सूक्ष्म कॉलम के निष्कर्षण के बाद, खोखले इंटीरियर प्रकार मैं कोलेजन के साथ लेपित है astrocyte आसंजन और गठबंधन बंडल गठन के लिए एक पर्यावरण स्वतंत्र की आपूर्ति (चित्र 1b -1) । बाद में, लुमेन मस्तिष्क cortical astrocytes प्रसव चूहा पिल्ले (चित्र 1b-2) से पृथक के साथ वरीयता प्राप्त है । दो आयामी (2d) संरेखण तकनीक है कि बिजली के खेतों, micropatterned खांचे के आवेदन पर निर्भर करते हैं, और extracellular मैट्रिक्स (ECM) प्रोटीन पैटर्न के विपरीत, रहने पाड़ तकनीक में astrocyte संरेखण आत्म विधानसभा पर निर्भर करता है ऐसे सब्सट्रेट वक्रता (कॉलम आईडी), कोशिका घनत्व, और कोलेजन एकाग्रता के रूप में नियंत्रणीय चर के अनुसार५०,५१,५२। astrocytes अनुबंध और कोलेजन remodel, और एक द्विध्रुवी प्राप्त, longitudinally-संरेखित आकृति विज्ञान के अनुरूप प्राकृतिक पाड़ों vivo में मनाया (चित्र 1b-3) । दरअसल, हम सक्रिय रूप से इन केबल का उपयोग कर रहे है के रूप में अच्छी तरह से क्षतिग्रस्त सीएनएस के प्रतिकूल वातावरण के माध्यम से axonal पुनर्जनन को सुविधाजनक बनाने के साथ ही स्थानांतरण अपरिपक्व न्यूरॉन्स के लक्षित मार्गदर्शन के लिए भौतिक सब्सट्रेट के रूप में संरचनाओं की तरह, विशेष रूप से स्तनधारी glial निशान (फिगर 1C). इस अनुच्छेद के astrocytic सूक्ष्म कॉलम, चरण इसके विपरीत और इम्यूनोफ्लोरेसेंस की छवियों की उंमीद cytoarchitecture के लिए विस्तृत निर्माण विधि पेश करेंगे, और वर्तमान सीमाओं और भविष्य के निर्देशों पर एक व्यापक चर्चा तकनीक.
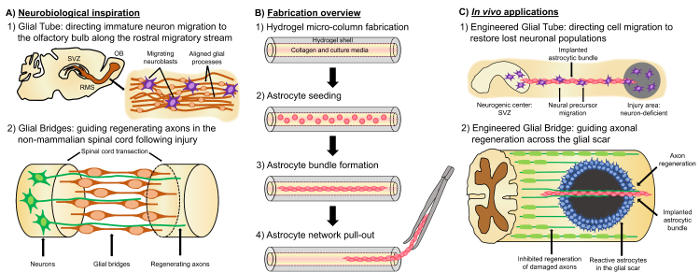
चित्रा 1: प्रेरणा, निर्माण प्रोटोकॉल, और गठबंधन Astrocytic नेटवर्क के लिए प्रस्तावित अनुप्रयोगों. (क) Neurobiological प्रेरणा: (१) तंत्रिकाजन्य subventricular जोन (SVZ) से Neuroblasts उद्भव longitudinally बल्ब (ओबी) की ओर निर्देशित प्रवास के लिए glial प्रवासी धारा (आरएमएस) में rostral संरेखित घ्राण ट्यूब का उपयोग; (2) उभयचर और मछली जैसे गैर स्तनधारियों एक glial पुल है कि एक घाव के सिरों को जोड़ता है के गठन के कारण भाग में तंत्रिका ऊतक क्षति के बाद पुनर्जनन बनाए रखने कर सकते है (जैसे transected रीढ़ की हड्डी) और के मार्गदर्शन के लिए एक पाड़ के रूप में कार्य करता है चूकने axons. (ख) निर्माण सिंहावलोकन: (1) एक माइक्रोन के निर्माण-आकार, खोखले hydrogel ECM के साथ लेपित लुमेन के साथ माइक्रो कॉलम, (2) प्राथमिक cortical astrocytes के सीडिंग प्रसव चूहा पिल्ले से पृथक, (3) longitudinally के आत्म-सभा उंमुख संस्कृति में बंडलों, और (4) भविष्य आरोपण अध्ययन के लिए झालर से बंडल के निष्कर्षण । (C) vivo अनुप्रयोगों में : (1) ये रहने वाले पाड़ तंत्रिकाजन्य केंद्रों से निर्देशित ंयूरॉन प्रवास के लिए इंजीनियर glial ट्यूबों के रूप में सेवा कर सकते है ंयूरॉन की कमी क्षेत्रों को फिर से आबाद; (२) अग्रणी axon मार्गदर्शन के विकासात्मक तंत्र की Recapitulation और गैर-स्तनधारियों में glial पुलों के पुनर्उत्पादक तंत्र के राज्यसत्ता इन astrocytic पाड़ों को गैर-axon के पार स्वतंत्र पुनर्जनन को प्रत्यक्ष करने की क्षमता के साथ- स्तनधारी glial निशान के पर्यावरण । कृपया यहां क्लिक करें इस आंकड़े का एक बड़ा संस्करण को देखने के लिए ।
