Het centrale zenuwstelsel (CNS) heeft een beperkte capaciteit voor het tegengaan van het verlies en/of disfunctie van neuronen en axonale trajecten die gepaard gaan met aandoeningen zoals traumatisch hersenletsel (TBI), beroerte, ruggenmerg letsel (SCI) en neurodegeneratieve ziekte1 ,2,,3,,4,5. Neurogenese in het VNV is beperkt tot een beperkt aantal gebieden in de hersenen, belemmering voor het herstel van verloren neuronen6,7. Bovendien, is regeneratie van verloren axonale trajecten in het VNV onvoldoende als gevolg van het ontbreken van gerichte begeleiding, de aanwezigheid van uitgroei remmers en reactieve astrogliosis na beschadiging zenuwweefsel2,8, 9,10. Astrocyten hebben gewoonlijk diverse functies in het bijstaan van neuronen met ion homeostase, neurotransmitter klaring, synaps formatie en neurovasculaire koppeling11. Niettemin, na zelfs milde beschadiging van zenuwweefsel, astrocyten kunnen moleculaire, structurele en functionele wijzigingen ondergaan als ze de overgang naar een hypertrofische staat11. In reactie op de ernstige neurotrauma, deze veranderingen leiden tot de vorming van een litteken met een bijschaduw met migreren reactieve astrocyten en een laesie kern waarin leukocyten gelekt uit de gescheurde bloed – hersenbarrière (BBB), microglia, oligodendrocyten en fibroblasten11,12,13. Deze reactieve astrocyten bereiken een morfologie van draadvormige, ongeorganiseerd processen en vertonen verhoogde expressie van tussenliggende gloeidraad eiwitten en chondroitin sulfaat-proteoglycans (CSPGs), die een belemmering vormen voor neurale regeneratie12. Hoewel het gliale litteken aanvankelijk helpt BBB integriteit herstellen en voorkomen van transmissie van de ontstekingsreactie naar het omliggende gezonde weefsel, dient het als een fysische en biochemische barrière tegen axon regeneratie12,14 ,15,16. Bijvoorbeeld, axonen die het gliale litteken optreden weergeven bolvormige Dystrofe groei kegels en onvolgroeide groei12. Bovendien belemmert de desorganisatie van astrocytic processen na blessure de uitbreiding van het regenereren van axonen17. Het resultaat van deze remmende eigenschappen komt ook tot uiting in de vaak-permanente lichamelijke en neurologische waardeverminderingen op die patiënten na ernstige neurotrauma lijden, met inbegrip van TBI en SCI.
Ongeacht de extrinsieke uitdagingen functionele regeneratie in het VNV, zijn axonen aangetoond te beschikken over een intrinsieke vermogen om te regenereren. Bijvoorbeeld, suggereert de dynamische aard van de kegels Dystrofe groei in contact met het gliale litteken dat deze uitgangen behouden hun capaciteit uit te breiden van12. Dus, het is van mening dat een belangrijke belemmering voor axonale uitgroei de remmende omgeving van de CNS na letsel en dat een meer tolerante omgeving is via vermindering van gliale littekens en/of verstrekken van regeneratieve bruggen over het litteken zou voordelige. Inderdaad, de vorige studies hebben aangetoond dat CNS neuronen aankonden axonen door middel van een laesie met behulp van perifere zenuwen transplantaten als bruggen, die een gunstiger omgeving voor axon regeneratie12,18, uit te breiden 19. Verschillende andere strategieën zijn uitgeoefend om te exploiteren deze rudimentair regeneratief vermogen. Bijvoorbeeld, heeft manipulatie van cel groei signaalroutes in verschillende modellen van de schade geleid tot axonale regeneratie en gliale scar vermindering10,20,21. Bovendien, hebben studies aangetoond dat behandeling met chondroitinase ABC, die de meerderheid van de suiker-ketens cleaves in CSPGs, de remmende werking van CSPGs uitgescheiden door reactieve astrocyten22 vermindert. Ondanks de bemoedigende resultaten, deze benaderingen bieden geen regie begeleiding van groei kegels, die kan leiden tot afwijkende regeneratie12, en ook geen rekening met het verlies aan neuronen. Cel-gebaseerde benaderingen hebben gebruikt in pogingen om te overwinnen van de gevolgen van de gliale litteken en om aan te vullen verloren cellen, met name neuronen. Sommige groepen hebben dedifferentiated reactieve astrocyten in neuronen, terwijl anderen hebben neurale voorlopercellen getransplanteerd in CNS laesies repopulate de schade-gebied en het bevorderen van axon regeneratie23,24, 25. stamceltransplantatie alleen wordt echter beperkt door lage overlevingskansen, slechte integratie en het bescheiden vasthouden in de beschadigde weefsel5. Bovendien, deze cel-gebaseerde strategieën niet herstellen van interlokale axonale traktaten, vooral op een gecontroleerde manier. Daarom, biomaterialen in combinatie met andere benaderingen worden onderzocht als leverende voertuigen voor diverse neurale en voorlopercellen en groei factoren26. Biomaterial-gebaseerde benaderingen zijn voorzien van een hoge mate van Ontwerpcontrole voor de productie van constructies die na te de specifieke fysieke, haptotaxic bootsen, en chemotaxic signalen aanwezig is in de driedimensionale (3D)-communicatie van de doel host weefsel27, 28,29,30,31,32,33,34. Reproductie van deze Milieusignalen is primordiaal voor getransplanteerde cellen te presenteren native-achtige morfologie, proliferatie, migratie en signalering, onder andere neurobiologische kenmerken29. Ondanks deze gunstige eigenschappen is vooruitgang dan traditionele cel geplaatste biomaterial steigers moeten tegelijkertijd bevorderen van gestuurde interlokale axonale regeneratie en vervanging van verloren neuronen.
“Levende steigers”, die te onderscheiden van andere cel-gebaseerde benaderingen als gevolg van de aanwezigheid van levende neurale cellen met een voorgevormde cytoarchitecture die native neuroanatomie emuleert een veelbelovende alternatieve aanpak is gebaseerd op zenuwweefsel ontworpen en/of Developmental mechanismen om gerichte vervanging, wederopbouw en herstel van de neurale circuits4,35. Overwegingen bij het ontwerp van levende steigers zijn de fenotypes en de bronnen van neurale cellen, evenals de mechanische/fysische eigenschappen en de biochemische signalen gedicteerd door de samenstelling van de begeleidende biomaterialen35. Na fabricage in vitro, deze levende steigers kunnen worden geïmplanteerd in vivo aan de huidige cel-celadhesie-moleculen en Chemotactische en neurotrophic signalen neurale cel migratie en axon uitgroei afhankelijk van de status actief worden gereguleerd en progressie van regeneratieve processen35. Gliale cellen kunnen dienen als basis voor de gemanipuleerde cytoarchitecture van levende steigers, aangezien deze cellen verschillende ontwikkelings mechanismen in vivo bemiddelen. Tijdens de ontwikkeling van de hersenen rekenen nieuwe neuronen op basale processen met radiale glia verlengd van de ventriculaire zone naar de ontwikkelingslanden corticale plaat als levende steigers voor gestuurde migratie36,37. Anderzijds uitbreiding van groei kegels zijn getoond om te oriënteren zich door het aftasten van de aantrekkelijke en afstotend signalen ontlokte door markering gliale cellen, en zogenaamde “baanbrekende” axonen worden voorgesteld om de juiste doelstellingen bereiken door uitbreiding langs vooraf gedessineerde gliale steigers35,38,39. Gliale cellen zijn dus nodig voor de begeleiding van baanbrekende axonen, die later als axon gebaseerde dienen “levende steigers” om de projectie van “volger” axonen. Bovendien groei glia-gemedieerde mechanismen te volharden postnatale stadium manifesteren, zoals neuroblasten de rostraal migratoire stroom (RMS volgen) om te navigeren van de subventriculaire zone (SVZ), een van de weinige resterende gebieden van neurogenese in de volwassen hersenen, is aangetoond dat de bulbus olfactorius (OB)40. Deze neuroblasten in de RMS migreren binnen de gliale buis (figuur 1A-1), die bestaat uit de lengterichting uitgelijnde astrocytic processen, via directe cel verklevingen en een gelokaliseerde oplosbare factoren37, 41. ten slotte terwijl CNS schade in zoogdieren oorzaken verstoord astrocytic proces regeling vormen een gliale litteken dat fysiek axonale regeneratie17 belemmert, veel bij niet-zoogdieren systemen ontbreken de vorming van een schadelijk gliale litteken. Daarentegen handhaven gliacellen bij niet-zoogdieren soorten meer georganiseerd, patronen die worden gebruikt als gidsen door de benadeelde regio17,42,43uitgelijnd. Bijvoorbeeld, worden bij niet-zoogdieren SCI modellen, axonen getoond om te groeien in nauwe samenwerking met gliale bruggen oversteken van de laesie, suggereren een belangrijke rol voor georganiseerde gliale steigers als substraten axonale regeneratie en functionele herstel (te vergemakkelijken Figuur 1A -2) 42 , 44 , 45. recapitulatie van de neuroanatomische functies en de ontwikkelings/regeneratieve mechanismen zoals hierboven beschreven kan het opleveren van een nieuwe klasse van gemanipuleerde gliale gebaseerde levende steigers die gelijktijdig onvolwassen neuronale migratie kan rijden en axonale vastloper door anders niet-tolerante omgevingen, waardoor potentieel afzwakking van de gevolgen van neuronale en axon tract degeneratie geassocieerd met CNS letsel en ziekte.
Onze onderzoeksgroep heeft eerder ontworpen meerdere soorten levende steigers voor wederopbouw en herstel van axonale traktaten in het centraal zenuwstelsel en het perifere zenuwstelsel (PNS) via micro-weefsel ontworpen neurale netwerken (micro-TENNs) en weefsel gemanipuleerde zenuw transplantaties (TENGs), respectievelijk27,46,47,48. Beide strategieën zijn inherent gebaseerd op het biomimicry. Micro-TENNs zijn anatomisch-geïnspireerde structuren ontworpen ter vervanging van structureel en functioneel axonale traktaten verbinden verschillende neuronale populaties van de hersenen. TENGs exploiteren de ontwikkelingstoxiciteit mechanisme van axon-vergemakkelijkt axonale regeneratie, wordt geïllustreerd door “volger” axon groei langs “pionier” axonen, om gerichte host axonale regeneratie35,46,48. We onlangs gekapitaliseerd op de veelzijdigheid van de levende steiger techniek met behulp van een soortgelijke regeling omhulsel als micro-TENNs en op zoek naar inspiratie uit de glia gebaseerde mechanismen aanwezig in de gehele ontwikkeling. Hier, ontwikkelde we constructies bestaande uit uitgelijnde astrocytic bundels verspreid over de collagene lumen van een hydrogel micro-kolom49. Deze astrocytic levende steigers zijn ontwikkeld door het eerste vullen een vergadering van de capillaire buis-acupunctuur-naald met vloeibare agarose als u wilt maken van een holle cilindrische hydrogel met een buitendiameter (OD) en de binnendiameter (ID) overeenkomt met de diameter van de buis en needle, respectievelijk. Na agarose gelering en extractie van de hydrogel micro-kolom van de capillaire buis, de holle interieur is bekleed met type I collageen leveren een omgeving tolerant voor Astrocyt hechting en uitgelijnd bundel vorming (figuur 1B -1). Daarna is de lumen bezaaid met cerebrale corticale astrocyten geïsoleerd van postnatale rat pups (figuur 1B-2). In tegenstelling tot tweedimensionale (2D) uitlijning technieken die afhankelijk zijn van de toepassing van elektrische velden, micropatterned groeven en extracellulaire matrix (ECM) eiwitten patronen, Astrocyt uitlijning in het levende schavot techniek is gebaseerd op zelf-assemblage volgens controleerbare variabelen zoals substraat kromming (kolom-ID), celdichtheid en collageen concentratie50,51,52. De astrocyten contract remodelleren van de collageen en verwerven een bipolaire, overlangs uitgelijnde morfologie analoog aan de natuurlijke steigers waargenomen in vivo (figuur 1B-3). Inderdaad, we actief het gebruik van deze kabel-achtige structuren nastreven als fysieke substraten voor gerichte begeleiding van onrijpe neuronen migreren, alsmede het vergemakkelijken van axonale regeneratie via het ongunstige milieu van het beschadigde centraal zenuwstelsel, met name de zoogdieren gliale litteken (Figuur 1 c). Dit artikel zal presenteren de gedetailleerde fabricage methode voor de astrocytic micro-kolommen, fase contrast en immunofluorescentie beelden van de verwachte cytoarchitecture en een uitgebreide discussie over de huidige beperkingen en toekomstige richtingen van de techniek.
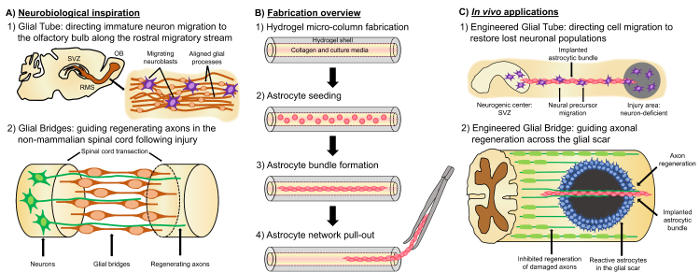
Figuur 1: inspiratie, Fabrication Protocol en voorgestelde toepassingen voor de uitgelijnde Astrocytic netwerken. (A) neurobiologische inspiratie: (1) neuroblasten die afkomstig zijn uit de neurogene subventriculaire zone (SVZ) gebruiken de lengterichting uitgelijnde gliale buis in de rostraal migratoire stroom (RMS) voor de geleide overgang naar de bulbus olfactorius (OB); (2) niet-zoogdieren zoals amfibieën en vis aankunnen regeneratie na zenuwweefsel beschadigen gedeeltelijk als gevolg van de vorming van een gliale brug verbindt de uiteinden van een laesie (bijvoorbeeld verbeelde ruggenmerg) en fungeert als een steiger voor de begeleiding van regenereren axonen. (B) fabricage overzicht: (1) de bouw van een micron-sized, holle hydrogel micro-kolom met de lumen bekleed met ECM, (2) zaaien van primaire corticale astrocyten geïsoleerd van postnatale rat pups, (3) zelf-assemblage van de lengterichting georiënteerde bundels in cultuur, en (4) extractie van de bundel van het omhulsel van de biomaterial voor toekomstige implantatie studies. (C) In vivo toepassingen: (1) deze levende steigers kunnen dienen als gemanipuleerde gliale buizen voor gestuurde neuron migratie van neurogene centra aan repopulate neuron-deficiënte regio’s; (2) de recapitulatie van de ontwikkelingstoxiciteit mechanisme van baanbrekende axon begeleiding en de regeneratieve mechanisme van gliale bruggen bij niet-zoogdieren kan deze astrocytic steigers met het vermogen om directe axon regeneratie over de niet-tolerante begiftigen omgeving van de zoogdieren gliale litteken. Klik hier voor een grotere versie van dit cijfer.
