Merkezi sinir sistemi (MSS) kaybı ve/veya nöronlar ve travmatik beyin hasarı (TBY), gibi koşullar eşlik aksonal yolları disfonksiyon karşı koymak için sınırlı bir kapasiteye sahip inme, omurilik yaralanma (SCI) ve nörodejeneratif hastalık1 ,2,3,4,5. MSS neurogenesis beyin, kayıp nöronlar6,7restorasyonu engelleyici alanda sınırlı sayıda sınırlıdır. Ayrıca, MSS kayıp aksonal yollar rejenerasyon yönlendirilmiş rehberlik eksikliği, akıbet inhibitörleri ve reaktif astrogliosis sinir dokusu2,8‘ ehasar takip varlığı nedeniyle yetersiz olduğunu, 9,10. Astrocytes genellikle nöronlar iyon Homeostazı, nörotransmitter gümrükleme, sinaps oluşumu ve11kaplin nörovasküler yardımcı çeşitli işlevlere sahiptir. Yine de, onlar Hipertrofik devlet11‘ e geçiş olarak sinir dokusu hatta hafif hasar, moleküler, yapısal ve fonksiyonel değişiklikleri astrocytes meydana gelebilir. Şiddetli neurotrauma yanıt olarak, bu bir yara oluşumuna geçirme reaktif astrocytes ve parçalanmış kan – beyin bariyerini (BBB), microglia sızan lökosit içerir bir lezyon çekirdek içeren bir penumbra ile değiştirilmesinden, oligodendrocytes ve fibroblastlar11,12,13. Bu reaktif astrocytes ipliksi, dağınık süreçlerin bir Morfoloji ulaşmak ve ara filaman proteinler ve nöral rejenerasyon12engel kondroitin sülfat Proteoglikanlar (CSPGs), artan ifadesi sergilemek. Gliyal yara başlangıçta BBB bütünlüğünün ve sağlıklı dokuyu çevreleyen inflamatuar yanıtın iletimini önlemek yardımcı olur olsa bile, bu axon rejenerasyon12,14 karşı fiziksel ve biyokimyasal bariyer olarak hizmet vermektedir ,15,16. Örneğin, gliyal yara karşılaşma aksonlar soğanlı distrofik büyüme koniler görüntülemek ve büyüme12bodur. Ayrıca, astrositik süreçleri dağınıklığı yaralanma sonra aksonlar17Yenileyici uzantısı engellemektedir. Bu inhibitör özellikleri sonucu hastaların Tby ve bilimler de dahil olmak üzere ağır neurotrauma sonra acı kez kalıcı fiziksel ve nörolojik bozukluklar içinde kendini gösteriyor
MSS fonksiyonel rejenerasyon karşı karşıya dışsal sorunları ne olursa olsun, akson yeniden oluşturmak için içsel bir yeteneğine sahip gösterilmiştir. Örneğin, gliyal yara ile temas distrofik büyüme koniler dinamik doğası bu sonlar12genişletmek için kapasitelerini korur düşündürmektedir. Sonuç olarak, bu aksonlar yeniden büyüme için bir ana engel sonrası yaralanma CNS ve gliyal skarlasma ve/veya yara izi arasında rejeneratif köprüler olurdu sağlayan azaltılması yoluyla fazla izin veren bir ortam sağlayan inhibitör bir ortamdır inanılıyor avantajlı. Gerçekten de, önceki çalışmalarda CNS nöronlar aksonlar periferik sinir grefti axon rejenerasyon12,18için daha uygun bir ortam mevcut köprüler olarak kullanarak bir lezyon üzerinden uzanan yetenekli olduğunu göstermiştir, 19. Çeşitli stratejiler tüylerinin bu rejeneratif kapasite yararlanma takip edilmiştir. Örneğin, hücre büyüme sinyal yolları çeşitli yaralanma modellerinde manipülasyon aksonal rejenerasyon ve gliyal yara izi azaltma10,20,21sonuçlandı. Ayrıca, çalışmalar chondroitinase şeker zincirleri çoğunluğu CSPGs içinde cleaves, ABC ile tedavi CSPGs reaktif astrocytes22tarafından salgılanan inhibitör etkisini azaltır göstermiştir. Rağmen cesaret verici sonuçlar, bu yaklaşımların potansiyel anormal rejenerasyon12‘ neden olabilir ve aynı zamanda nöronların kaybı için hesap yapmak büyüme koniler rehberliğinde yönetmen sağlamaz. Hücre tabanlı yaklaşımlar girişimleri gliyal yara etkilerini aşmak için ve kayıp hücreleri, özellikle sinir hücreleri doldurmak için kullanılmıştır. Diğerleri yaralanma alanı yeniden doldurmanız ve akson rejenerasyon23,24, teşvik CNS lezyonları nöral progenitör hücre nakli sırasında bazı gruplar reaktif astrocytes nöronlar dediferansiye 25. ancak, kök hücre transplantasyonu tek başına düşük sağkalım oranları, zavallı entegrasyon ve hasarlı doku5mütevazı saklama ile sınırlıdır. Ayrıca, hücre tabanlı strateji de uzun mesafe aksonal yolları, özellikle denetimli bir biçimde geri yüklemek başarısız. Bu nedenle, biyomalzemeler diğer yaklaşımlar ile birlikte teslim araçlar için çeşitli sinir incelenmiştir ve progenitor hücreler ve büyüme faktörleri26. Biomaterial tabanlı yaklaşımlar tasarım denetimi belirli fiziksel, haptotaxic taklit yapıları üretmek için yüksek derecede özelliği ve hedef ana bilgisayar doku27üç boyutlu (3D) microenvironment içinde chemotaxic ipuçları sunmak, 28,29,30,31,32,33,34. Bu çevre sinyalleri çoğaltılması yerli benzeri Morfoloji, nükleer silahların yayılmasına karşı geçiş ve, sinyal29arasında diğer nörobiyolojik özellikleri sunmak nakledilen hücreler için her şeyden önemlidir. Bu avantajlı özellikleri rağmen geleneksel hücre tohumlari biomaterial iskele ötesinde gelişme aynı anda yönettiği uzun mesafe aksonal rejenerasyon teşvik ve kayıp nöronlar değiştirmek için gereklidir.
Alternatif bir yaklaşım sinir dokusu üzerinde dayanır umut verici bir “yaşam sinir hücreleri ile yerel Nöroanatomi öykünür bir ön şekillendirilmesi cytoarchitecture varlığı nedeniyle diğer hücre tabanlı yaklaşımlar farklıdır yaşam iskele”, mühendislik ve/veya hedeflenen değiştirme, yeniden yapılanma ve yeniden nöro çevrim4,35oluşturulmasını kolaylaştırmak için gelişimsel mekanizmaları. Yaşam iskele tasarımı için önemli noktalar fenotipleri ve sinir hücreleri gibi mekanik/fiziksel özellikleri kaynakları ve biyokimyasal sinyalleri herhangi bir eşlik eden Biyomalzeme35kompozisyon tarafından dikte. İmalat vitrosonra bu yaşam iskele implante vivo içinde mevcut hücre adezyon molekülleri ve kemotaktik olabilir ve Nörotrofik sinyalleri aktif sinir hücre göç ve akson akıbet durumuna bağlı olarak düzenleyen ve rejeneratif ilerlemesini35işler. Bu hücreler çeşitli gelişimsel mekanizmaları vivo içindearacılık beri gliyal hücreler yaşam iskele mühendislik cytoarchitecture için bir temel olarak hizmet verebilir. Beyin gelişimi sırasında Bazal süreçleri yaşayan iskele yönlendirilmiş geçiş36,37için olarak gelişmekte olan kortikal plaka doğru ventrikül bölgeden Radyal glia tarafından genişletilmiş yeni nöronlar güveniyor. Ayrıca, kendilerini çekici ve itici sinyalleri gliyal Işaret hücreleri tarafından elde edildi algılama tarafından yönlendirmek için gösterilen ve sözde “aksonlar öncü” koniler büyüme uzanan önerilir boyunca önceden desenli gliyal genişleterek doğru hedeflere ulaşmak için 35,38,39iskele. Böylece, gliyal hücreler daha sonra olarak axon tabanlı hizmet aksonlar öncülük etmek için sağlanan bilgilerde açıklanan gerekli olan “yaşam iskele” projeksiyon “takipçisi” akson ve yönlendirmek için. Ayrıca, büyüme glia aracılıklı mekanizmalar olarak rostral göçmen akışı (RMS) bölgesinden subventricular (SVZ), bir yetişkin beyninin neurogenesis birkaç diğer alanlarında gezinmek için neuroblasts takip postnatally, kalıcı olması için gösterilmiştir olfaktör ampul (OB)40. Bu neuroblasts RMS boyuna hizalanmış astrositik oluşum, doğrudan hücre-hücre yapışıklıklar yolu ile oluşan ve çözünür faktörler37, lokalize gliyal tüp içinde (şekil 1A-1), göç 41. memeliler nedenleri CNS hasar fiziksel olarak aksonal rejenerasyon17engellemektedir gliyal bir yara izi oluşturan astrositik işlem düzenleme kesintiye iken, son olarak, birçok memeli sistemi zararlı gliyal yara oluşumu eksikliği. Daha doğrusu, Sigara memeli türlerinin gliyal hücreler daha organize, yaralı bölge17,42,43kılavuzları olarak kullanılan desenler hizalı korumak. Örneğin, memeli SCI modellerinde, gliyal köprüler yüzeylerde aksonal rejenerasyon ve fonksiyonel iyileşme (kolaylaştırıcı olarak organize gliyal iskele için önemli bir rol düşündüren lezyon, geçiş ile yakın ilişki içinde büyümeye aksonlar gösterilir Şekil 1A -2) 42 , 44 , 45. tekrarlama nöroanatomik özellikleri ve yukarıda açıklanan gelişimsel/rejeneratif mekanizmaları aynı anda olgunlaşmamış nöronal göç sürebilirim mühendislik gliyal tabanlı yaşam iskele yeni bir sınıf verim ve aksonal Bunun aksi halde keyfi ortamlar, böylece potansiyel olarak nöronal etkilerini Azaltıcı aracılığıyla ve akson yolu dejenerasyonu CNS yaralanma ve hastalık ile ilgili.
Bizim araştırma grubu daha önce yaşayan iskele yeniden inşası için birden çok türde Dizayn ve rejenerasyon aksonal yollarini MSS ve mikro-doku ile periferik sinir sistemi (PNS) sinir ağları (mikro-TENNs) ve doku Mühendisliği sinir grefti (TENGs), sırasıyla27,46,47,48mühendislik. Hem stratejileri doğal olarak Biyomimikri üzerinde temel alır. Mikro-TENNs yapısal ve işlevsel olarak beynin farklı nöron popülasyonları bağlanma aksonal yolları değiştirmek için tasarlanmış anatomik olarak ilham yapılardır. TENGs gelişimsel mekanizması “takipçisi” axon büyüme hedeflenen ana bilgisayar aksonal rejenerasyon35,46,48elde etmek için “öncü” akson boyunca örneği axon kolaylaştırdı aksonal rejenerasyon yararlanmak. Biz son zamanlarda yaşayan iskele çok yönlülük büyük harfle tekniği benzer bir encasement şeması mikro-TENNs ve glia tabanlı mekanizmaları ilham arayan mevcut geliştirme. Burada, kolajen Lümen hidrojel mikro-sütun49kapsayan hizalanmış astrositik demetleri oluşan yapıları geliştirdik. Bu astrositik yaşam iskele çapları için karşılık gelen bir içi boş silindirik hidrojel bir dış çap (OD) ve iç çapı (ID) ile oluşturmak için bir kılcal tüp-akupunktur iğne derleme sıvı özel ile doldurarak geliştirilir tüp ve iğne, anılan sıraya göre. Özel jelleşme ve hidrojel mikro-sütun çıkarma kılcal tüp, içi boş iç kısmıdır kaplı türü ile ben astrocyte yapışma için izin veren bir ortam sağlamak için kollajen ve paket oluşumu (şekil 1B uyumlu -1). Daha sonra lumen postnatal fare pups (şekil 1B-2) izole serebral kortikal astrocytes ile seribaşı. Elektrik alanları, micropatterned oluklar ve hücre dışı matriks uygulanması (ECM) protein desenlendirme, itimat iki boyutlu (2D) hizalama teknikleri astrocyte uyum içinde yaşayan iskele aksine tekniği üzerinde kendinden montajlı dayanıyor substrat eğriliği (sütun Kımlığı), hücre yoğunluğu ve kollajen konsantrasyonu50,51,52gibi kontrol değişkenleri göre. Astrocytes sözleşme ve kollajen yeni model ve iki kutuplu, boyuna hizalı Morfoloji vivo içinde (şekil 1B-3) gözlenen doğal iskele benzer elde etmek. Nitekim, biz aktif olarak bu kablo benzeri yapıların kullanımını hedeflenen rehberlik olgunlaşmamış nöronlar geçiş hem de özellikle bozuk MSS olumsuz ortamı üzerinden aksonal rejenerasyon kolaylaştırmak için fiziksel yüzeyler olarak devam ediyor memeli gliyal yara (şekil 1 c). Bu makalede, astrositik mikro-sütunlar için detaylı imalat yöntemi mevcut beklenen cytoarchitecture ve mevcut kısıtlamaları üzerinde kapsamlı bir tartışma kontrast ve ayirt görüntülerini ve gelecekteki yönleri faz tekniği.
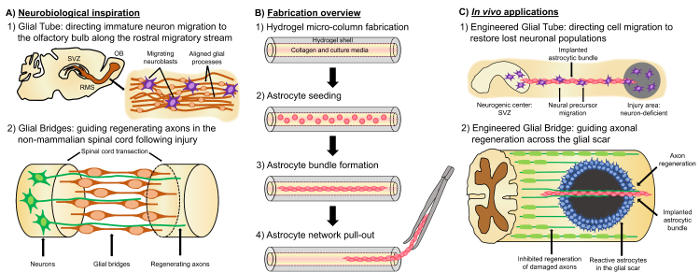
Şekil 1: ilham, imalat Protokolü ve hizalanmış astrositik ağlar için Önerilen uygulamalar. (A)nörobiyolojik ilham: (1) Neuroblasts Nörojenik subventricular bölgesinden (SVZ) kaynaklı boyuna hizalanmış gliyal tüp rostral göçmen akışı (RMS) olfaktör ampul (OB); doğru yönlendirilmiş geçiş için kullanmak (2) sonra sinir dokusuna zarar kısmen-bir lezyon (örneğin kopuk spinal kord) ucuna bağlanan ve rehberlik için bir iskele olarak hizmet veren bir gliyal köprü oluşumu nedeniyle kurbağa ve balık gibi sigara-memeliler rejenerasyon ayakta olabilir akson yeniden oluşturuluyor. (B) imalat genel bakış: mikron büyüklüğünde, içi boş hidrojel mikro-sütun ECM ile kaplı Lümen ile inşaat (1) (2) postnatal fare pups (3) kendinden montajlı, boyuna yönelik izole birincil kortikal astrocytes, tohum Kültür ve paket (4) çekme–dan biomaterial mantolama gelecekteki implantasyon Etütler demetleri. (C) In vivo uygulamaları: (1) bu yaşam iskele mühendislik gliyal tüpler için yönlendirilmiş nöron göç–dan nöron eksikliği bölgeleri; yeniden oluşturulacaktır için Nörojenik merkezleri olarak hizmet verebilir (2) tekrarlama axon rehberlik öncülük etmek gelişimsel mekanizması ve sigara-memelilerde gliyal köprüler rejeneratif mekanizması ile axon rejenerasyon sigara-keyfi arasında yönlendirmek için kapasite astrositik bu iskele bağışlamak memeli gliyal yara ortamı. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.
