Det centrala nervsystemet (CNS) har begränsad förmåga att motverka den förlust eller dysfunktion av nervceller och axonal vägar som åtföljer villkor såsom traumatisk hjärnskada (TBI), stroke, ryggmärgen skadan (SCI) och neurodegenerativa sjukdomen1 ,2,3,4,5. Neurogenes i CNS är begränsad till ett begränsat antal områden i hjärnan, hämmar återställandet av förlorade nervceller6,7. Dessutom är regenerering av förlorade axonal vägar i CNS otillräcklig på grund av bristen på riktad vägledning, förekomsten av utväxt hämmare och reaktiva astrogliosis efter skada till nervvävnad2,8, 9,10. Astrocyterna har vanligtvis olika funktioner hjälpa nervceller med ion homeostas, signalsubstansen clearance, synaps bildas och neurovaskulära koppling11. Dock efter även lindriga skador till nervvävnad, kan astrocyter genomgå molekylär, strukturella och funktionella förändringar som de övergången till en hypertrofisk stat11. Svar på svåra neurotrauma resultera ändringarna i bildandet av ett ärr med en penumbra innehållande migrerar reaktiva astrocyter och en lesion kärna som innehåller leukocyter som läckt ut från en brusten blod – hjärnbarriären (BBB), mikroglia, oligodendrocyter och fibroblaster11,12,13. Dessa reaktiva astrocyter uppnå en morfologi av trådformiga, oorganiserad processer och uppvisar ökat uttryck av mellanliggande glödtråden proteiner och chondroitin sulfate proteoglykaner (CSPGs), som hindrar neurala regenerering12. Även om gliaceller ärret hjälper först återställa BBB integritet och undvika överföring av den inflammatoriska reaktionen till omkringliggande frisk vävnad, fungerar det som en fysiska och biokemiska barriär mot axon regenerering12,14 ,15,16. Exempelvis axoner som möter gliaceller ärret Visa uppsvällda dystrofa tillväxt kottar och hämmad tillväxt12. Dessutom hindrar desorganisation av astrocytic processer efter skada utvidgning av regenererande axoner17. Resultatet av dessa hämmande egenskaper manifesteras i de ofta permanenta fysiska och neurologiska funktionsnedsättningar som patienter lider efter svår neurotrauma, inklusive TBI och SCI.
Oavsett de yttre utmaningarna funktionell förnyelse i CNS, har axoner visat sig ha en inneboende förmåga att regenerera. Exempelvis antyder den dynamiska karaktären av dystrofa tillväxt kottarna i kontakt med gliaceller ärret att dessa ändelser behåller sin kapacitet att utöka12. Följaktligen, man tror att ett främsta hinder för axonal re-tillväxt är hämmande miljön efter skada CNS och som ger en mer tillåtande miljö via minska gliaceller ärrbildning och/eller tillhandahålla regenerativ broar över ärret skulle vara fördelaktiga. Ja, tidigare studier har visat att CNS nervceller kunde utvidga axoner genom en lesion med perifer nerv ympkvistar som broar, som medför en mer gynnsam miljö för axon regenerering12,18, 19. Flera andra strategier har bedrivits för att utnyttja denna rudimentära förnyelseförmåga. Till exempel har manipulation av cell tillväxt signalvägar i olika skada modeller resulterat i axonal regenerering och gliaceller scar minskning10,20,21. Studier har dessutom visat att behandling med chondroitinase ABC, som klyver majoriteten av sockerkedjor i CSPGs, minskar den hämmande effekten av CSPGs utsöndras av reaktiva astrocyter22. Trots de uppmuntrande resultat, dessa metoder ger inte riktat vägledningen av tillväxt kottar, vilket potentiellt kan resultera i aberrant regeneration12, och också inte redovisa förlust av nervceller. Cellbaserade metoder har använts i försök att övervinna effekterna av gliaceller ärret och att återanskaffa förlorade celler, särskilt nervceller. Vissa grupper har dedifferentiated reaktiv astrocyter till nervceller, medan andra har transplanterats neurala stamceller in i CNS-lesioner att återbefolka området skada och främja axon regenerering23,24, 25. stamcellstransplantation ensam är dock begränsad av låga överlevnaden, dålig integrering och blygsamma lagring i skadad vävnad5. Dessutom misslyckas dessa cellbaserade strategier att återställa långdistans axonal skrifter, särskilt på ett kontrollerat sätt. Därför biomaterial i kombination med andra metoder utforskas som leveransfordon för olika neurala och stamceller och tillväxt faktorer26. Biomaterial-baserade metoder har en hög grad av kontroll att producera konstruktioner som efterliknar den specifika fysiska, haptotaxic och chemotaxic cues presentera i den tredimensionella (3D) närmiljön för target host vävnad27, 28,29,30,31,32,33,34. Reproduktion av dessa Miljösignaler är avgörande för transplanterade celler presentera native morfologi, spridning, migration och signalering, bland andra neurobiologiska egenskaper29. Trots dessa fördelaktiga egenskaper krävs avancemang bortom traditionella cell seedade biomaterial ställningar att samtidigt främja riktad långdistans axonal regenerering och ersätta förlorade nervceller.
En lovande alternativa tillvägagångssätt bygger på nervvävnad konstruerad ”levande ställningar”, som är åtskilda från andra cell-baserat tillvägagångssätt på grund av levande neurala celler med en förformade cytoarchitecture som emulerar infödda neuroanatomi och/eller utvecklingsmässiga mekanismer för att underlätta riktade utbyte, återuppbyggnad och regenerering av neurala kretsar4,35. Överväganden för utformningen av levande ställningar inkluderar fenotyper och källor av neurala celler, samt mekanisk/fysiska egenskaperna och de biokemiska signalerna av sammansättningen av någon medföljande biomaterial35. Efter tillverkning i vitro, dessa levande ställningar kan vara implanterad i vivo till nuvarande cell-adhesionsmolekyler och kemotaktisk och neurotrofa signalerar att aktivt reglera neurala cellmigration och axon utväxt beroende på tillståndet och progression av regenerativ processer35. Gliaceller kan fungera som underlag för den bakåtkompilerade cytoarchitecture av levande ställningar eftersom dessa celler medla olika utvecklande mekanismer i vivo. Under hjärnans utveckling åberopa nya nervceller basala processer utvidgad genom radiella glia från ventrikulära zonen mot utveckla kortikala plattan som levande ställningar för riktad migration36,37. Dessutom sträcker sig tillväxt kottar är visat att orientera sig genom att känna attraktivt och motbjudande signaler som framkallas av gliaceller VÄGSTOLPE celler, och så kallade ”banbrytande” axoner föreslås för att nå de rätta målen genom att utvidga längs pre mönstrade gliaceller ställningar35,38,39. Gliaceller är således nödvändiga för vägledningen av banbrytande axoner, som senare tjäna som axon-baserade ”levande ställningar” direkt projektionen av ”efterföljare” axoner. Dessutom glia-medierad tillväxt mekanismer har visats kvarstå efter födseln, som neuroblasts följer rostralt flyttande strömmen (RMS) för att navigera från den subventrikulära zonen (SVZ), en av de få återstående områdena av neurogenes i den vuxna hjärnan, att den luktbulben (OB)40. Dessa neuroblasts i RMS migrera inom gliaceller röret (figur 1A-1), som består av longitudinellt-anpassade astrocytic processer, via direkt cell-cell sammanväxningar och lokaliserade lösliga faktorer37, 41. Slutligen, medan CNS skada i däggdjur orsakar stört astrocytic processen arrangemang bildar ett gliaceller ärr som fysiskt hindrar axonal regenerering17, många icke däggdjur system saknar bildandet av ett skadligt gliaceller ärr. Snarare, upprätthålla gliaceller icke däggdjur arter mer organiserad, justerad mönster som används som guider genom den skada regionen17,42,43. Exempelvis i icke däggdjur SCI modeller visas axoner att växa i nära samarbete med gliaceller broarna över lesionen, vilket tyder på en viktig roll för organiserade gliaceller ställningar som substrat underlätta axonal regenerering och funktionell återhämtning ( Figur 1A -2) 42 , 44 , 45. rekapitulation neuroanatomiska funktioner och de utvecklingsmässiga/regenerativa mekanismer som beskrivs ovan kan ge en ny klass av modifierade gliaceller-baserade levande ställningar som kan samtidigt driva omogna neuronala migration och axonal pathfinding genom annars icke-tillåtande miljöer, därmed potentiellt mildra effekterna av neuronala och axon tarmkanalen degeneration är associerad med CNS skada och sjukdom.
Vår forskargrupp har tidigare designat flera typer av levande ställningar för återuppbyggnad och regenerering av axonal skrifter i CNS och det perifera nervsystemet (PNS) via micro-vävnad konstruerade neurala nätverk (mikro-villkoren) och vävnad bakåtkompilerade nerv transplantat (TENGs), respektive27,46,47,48. Båda strategierna baseras sig på biomimicry. Micro-villkoren är anatomiskt-inspirerade strukturer för att strukturellt och funktionellt ersätta axonal skrifter ansluta distinkta neuronala populationer av hjärnan. TENGs utnyttja utvecklingsmässiga mekanismen av axon-stödda axonal regenerering, exemplifierat av ”efterföljare” axon tillväxt längs ”pioneer” axoner, att uppnå riktade värd axonal regenerering35,46,48. Vi nyligen balanserade på mångsidigheten hos levande schavotten tekniken använder ett liknande system för fodral som mikro-villkoren och söker inspiration från de glia-baserade mekanismerna presentera hela utveckling. Här utvecklade vi konstruktioner bestående justerad astrocytic buntar spanning kollagena lumen en hydrogel mikro-kolumn49. Dessa astrocytic levande ställningar är utvecklat av första fylla en kapillär tube-akupunktur nål församling med flytande agarosgelelektrofores för att skapa en ihålig cylindrisk hydrogel med ytterdiameter (OD) och innerdiameter (ID) motsvarar diametrarna på de röret och nål, respektive. Efter agaros gelation och utvinning av hydrogel mikro-kolumnen från kapillärröret, ihåliga interiören är belagda med typ I kollagen att tillhandahålla en miljö som är tillåtande för Astrocyten vidhäftning och justerad bunt bildandet (figur 1B -1). Efteråt, lumen är seedade med cerebral kortikal astrocyter isolerade från postnatal råttungar (figur 1B-2). Tvärtemot tvådimensionell (2D) justering tekniker som förlitar sig på tillämpningen av elektriska fält, micropatterned grooves och extracellulär matrix (ECM) protein mönstring, astrocyt justering i levande schavotten bygger teknik på självmontering enligt kontrollerbara variabler såsom substrat krökning (kolumnen ID), cell densiteten och kollagen koncentration50,51,52. Astrocyterna kontrakt och omskapa kollagen och förvärva en bipolär, longitudinellt-anpassade morfologi som är analoga med de naturliga ställningar som observerats i vivo (figur 1B-3). Faktiskt, vi bedriver aktivt användningen av dessa kabel-liknande strukturer som fysiska substrat för målinriktad vägledning av migrera omogna nervceller samt underlätta axonal regenerering genom den ogynnsamma miljön i skadade CNS, särskilt däggdjur gliaceller ärret (figur 1 c). Denna artikel kommer att presentera metoden detaljerade tillverkning för astrocytic mikro-kolumner, fas kontrast och immunofluorescens bilder av den förväntade cytoarchitecture och en omfattande diskussion om de nuvarande begränsningarna och framtida inriktningar av de teknik.
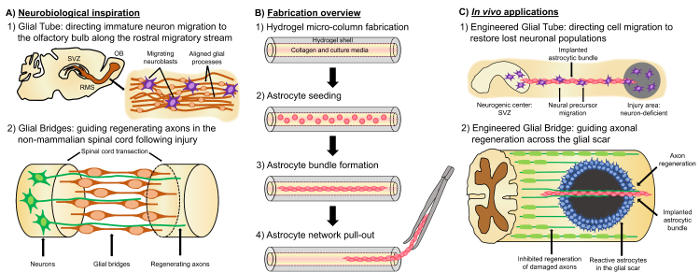
Figur 1: Inspiration, Fabrication protokoll och föreslagna program för de justerade Astrocytic nätverk. (A) neurobiologiska inspiration: (1) Neuroblasts med ursprung från neurogen subventrikulära zonen (SVZ) utnyttja längdriktningen justerad gliaceller röret i rostralt flyttande strömmen (RMS) för riktad övergång till luktbulben (OB); (2) icke-däggdjur såsom groddjur och fisk kan upprätthålla förnyelse efter nervvävnad skador delvis på grund av bildandet av en gliaceller bro som förbinder ändarna av en lesion (t.ex. transected ryggmärgen) och fungerar som en klätterställning för vägledningen av regenererande axoner. (B) tillverkning översikt: (1) byggandet av en micron stora, ihåliga hydrogel mikro-kolumn med lumen belagd med ECM, (2) sådd av primära kortikala astrocyter isolerade från postnatal råttungar, (3) självmontering av longitudinellt-orienterade buntar i kultur och (4) utvinning av bunten från den biomaterial fodral för framtida implantation studier. (C) In vivo program: (1) dessa levande ställningar kan tjäna som modifierade gliaceller rör för riktad neuron migrering från neurogen centra att återbefolka neuron-brist regioner. (2) rekapitulation av utvecklingstoxicitet mekanismen av banbrytande axon vägledning och regenerativ mekanismen av gliaceller broar i icke-däggdjur kan begåva dessa astrocytic ställningar med kapacitet att direkt axon regenerering över den icke-tillåtande miljön av däggdjur gliaceller ärr. Klicka här för att se en större version av denna siffra.
