植物関連微生物は、生物地球化学サイクル、バイオレメディエーション、気候変動の緩和、植物の成長と健康、生物的および非生物的ストレスに対する植物耐性において重要な役割を果たす。微生物は直接植物細胞壁の接触を介して間接的に化学分泌を介して1、2、3シグナリングの両方の植物と相互作用します。固着生物として、植物は病原体による感染に抵抗する直接的および間接的メカニズムを開発してきた。間接的な防御は、二次植物代謝物の生産と病原体4,5の侵入に対する拮抗生物の魅力を含むのに対し、直接防御は、構造的な防御および防御タンパク質の発現を含みます。植物由来の滲出液、分泌物、粘液、ムシゲル、および溶解物は、根圏の物理化学的特性を変化させて誘引または反発する宿主に向かって微生物6 。根分泌物の化学組成は種特異的であり、それによって、そのような化合物を認識することができる特定の微生物が根圏6で繁殖することを可能にする選択的フィルターとして役立つ。したがって、適合性の微生物種は、植物宿主1の有益性または有害性のいずれかに、それらの関連性を活性化および強化するように刺激され得る。
微生物および化学的暴露の大部分は、ルート構造と土壌エアインタフェース2、6、7、8で起こるからである。根圏における理解植物微生物相互作用は、植物生産性と生態系の機能を強化するための鍵でありますしかしながら、地下の植物 – 微生物相互作用および相反応答の試験は、その興味深いことに複雑かつ動的な性質、および厳密に制御可能な増殖条件下での天然根構造および植物形態を有する適切な実験モデルの欠如を含む。最も頻繁に研究植物病原の一つとして、 アグロバクテリウムは桜、リンゴ、ナシ、ブドウなどの農園芸重要性を持つ植物の広い範囲に感染し、9ローズ 。 アグロバクテリウムは理解植物-病原体相互作用のための重要なモデル生物であり、植物の形質転換および植物工学10、11、12、13、14における強力なツールです。
分子は、 アグロバクテリウムの相互作用が十分に数十年にわたって研究されている、およびアグロバクテリウム病原性の現在の理解は9広範であるプラント-F "> 11、15、16。 アグロバクテリウムの病原性は、主に17を感知する、植物由来の信号を感知その毒性プログラムおよび細胞間通信の細かい調節をもたらす、いわゆる定足数のその進化能力に起因する。ザアグロバクテリウムの病原性プログラムは根圏で利用可能ないくつかの信号によって調節され、2成分系、は、chvg / Iシステムとビラ/ Gシステムの2つのセットを含むている。根圏における酸性条件は、 は、chvg / I、 ビラ/ Gの転写を活性化します、およびvirE0 、 virE1 、 virH1 、 virH2 、およびVI型分泌系(T6SS) 18の遺伝子を含むアグロバクテリウムの病原性に関与するいくつかの他の遺伝子が含まれる。アセトシリンゴン(4'-ヒドロキシ-3 '、5 ' – ジメトキシアセトフェノン)、Vリン酸化シグナル伝達機構によるirA / G二成分系19 。その後、VirA / Gはvirレギュロン全体を活性化し、その結果、腫瘍誘導性(Ti)プラスミドからトランスファーDNA(T-DNA)と呼ばれる約20kbのバクテリアDNA断片を植物核16に移入して組み込む。 T-DNAは、植物ホルモンインドール-3-酢酸(IAA)( iaaMおよびiaaH )およびサイトカイニン( ipt )の合成に関与する遺伝子を保有し、一旦植物細胞で発現すると、これらの植物ホルモンが大量に産生される。これは、植物9、11、20のために慢性および復活問題であるクラウンゴール病として知られる異常な組織増殖および植物腫瘍発達、もたらします。 IAAは、 アグロバクテリウムビルレンスを抑制するために、またはアグロバクテリウムを減少させるために、サリチル酸およびγ-アミノ酪酸と一緒に作用する m個のクオラムセンシング(QS)17、21、22。この抑制に対抗するために、T-DNAはまた、 アグロバクテリウムの病原性を促進するために、 アグロバクテリウムクオラムセンシングを活性化し、病原体22,23のための栄養源となるオピンの生合成のための遺伝子を運びます。
アグロバクテリウム –プラント相互作用の全体的な深い理解と、植物宿主への得られたT-DNA転移にもかかわらず、相互作用の初期段階での複雑なシグナル伝達事象はあまり理解されていない。これは、部分的に、 アグロバクテリウム –植物シグナル伝達を調べるための従来のアプローチの限界に起因する。植物細胞懸濁液培養および人工部位特異的創傷は、分子植物 – 微生物相互作用を研究するために一般的に使用されている24 、。EF "> 26、27はしかし、細胞懸濁液は、一般的な植物の形態に欠け、特に、植物の懸濁細胞は、微生物の走化性および病原性28、29を活性化するために非常に重要なルート構造と根の滲出液を、持っていない植物の形態の維持。ルート構造が直接感染した植物組織30、31に誘導される植物防御関連遺伝子の検出結果として、サイト固有の感染を容易に人為的に負傷植物によって対処されてきた。しかし、人工的な創傷は、自然の中で病原体感染と大きく異なっています特に、創傷がジャスモン酸(JA)の蓄積をもたらし、天然植物のシグナル伝達と防御を全身的に妨害する26。また、合成化学物質は、人工的に植物宿主応答を誘導するために使用される26 。または病原体の病原性である。 植物体中の濃度を反映するような化学物質の補充が可能であるが、そのような補充は徐々に微生物28、32によって検知された走化性勾配を生成取り囲む根圏へ根滲出液の拡散を考慮していません。植物 – 微生物相互作用を研究する従来のアプローチの限界を考えると、得られたデータの精度と深さは妨げられ、制限され、従来のアプローチから生成された知識は植物に直接翻訳されない可能性がある。植物 – アグロバクテリウムシグナリングの多くの局面は、特に、疾患の症状がまだ発症していない初期の相互作用の段階では、完全には理解されていない。
従来のアプローチの限界を修正するために、この研究は、安価で、しっかりと制御可能で、柔軟な水耕性のc研究者が分子植物 – 微生物相互作用の初期段階で複雑なシグナル伝達経路と応答経路に深い洞察を得ることを可能にする。水耕栽培は広く植物栄養素、根の滲出液、成長条件、および植物33、34上の金属毒性の影響を研究するために使用されてきました。水耕栽培モデルには、小さな空間的要件、様々な植物組織の接近可能性、栄養素/環境条件の厳密な制御、害虫/病害の制御などのいくつかの利点があります。水耕栽培システムはまた、典型的には2〜3週間後に成長を制限する寒天/フィトアガルめっき技術と比較して、植物の生育を制限するものではない。重要なのは、全体のプラント構造物の維持管理は、微生物の走化性および病原性の誘導8、29のために必要な自然なルート分泌を促進します。システムの説明ここでのベッドは選択肢33、34よりも簡単かつ少ない労働集約的です。これは、より少ない部品を使用し、標準はさみ以外の工具を必要としません。それは、植物生長の強力な支持体としての金属メッシュ(ナイロン33とは対照的に)と、微生物増殖を助けるための振盪による無菌条件下での単純な通気方法を使用する。さらに、このシステムは、根の幅を制限することなく多様な植物種に適応する、植物成長をサポートするために様々なサイズの金属メッシュを使用することができる。
ここに提示されている水耕共培養系では、接種された細菌の増殖を支える有機化合物を植物の根が分泌する無菌水耕システムで植物を栽培する。この共培養システムでは、植物ホルモン、防御エリシター、または病原性誘導化学物質などの人工化学物質は補充されず、自然細胞を反映する植物 – 微生物相互作用中のホメオスタシスシグナル。この水耕共培養システムでは、同時に遺伝子、アグロバクテリウムによる感染時シロイヌナズナ COL-0根組織における発現、ならびにシロイヌナズナとの共培養時にアグロバクテリウム遺伝子の活性化を決定することが可能でした。このシステムは、 アグロバクテリウム ( Agrobacterium )との同時培養(感染)の際に、植物根へのアグロバクテリウムの付着および植物根の分泌のプロファイルを研究するのに適していることがさらに実証された( 図1 )。
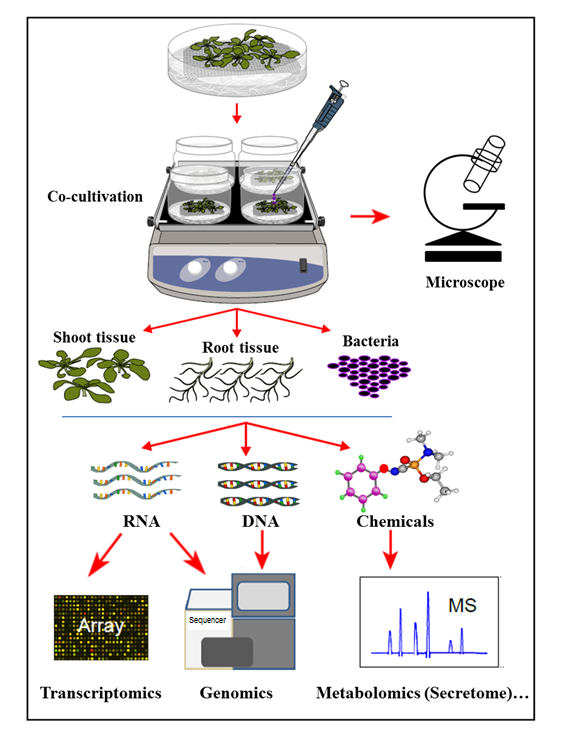
図1:サンプル分析を用いた水耕共培養系の概要。植物をメッシュの上に成長させる(メッシュの上に芽を入れる)が、水耕培地に浸漬され、その後、バクテリアfまたは共培養する。次いで、植物組織および細菌を分離して、同時に抽出および分析する。この数字は参考文献35から変更されています。
