Os micróbios associados à planta desempenham papéis importantes no ciclismo biogeoquimico, na biorremediação, na mitigação das alterações climáticas, no crescimento e na saúde das plantas e na tolerância das plantas aos estresses bióticos e abióticos. Os microorganismos interagem com as plantas diretamente através do contato da parede celular da planta e indiretamente através de secreção química e sinalização 1 , 2 , 3 . Como organismos sésseis, as plantas desenvolveram mecanismos diretos e indiretos para resistir à infecção por agentes patogênicos. As defesas diretas incluem defesas estruturais e a expressão de proteínas de defesa, enquanto que as defesas indiretas incluem a produção secundária de metabólitos das plantas e a atração de organismos antagônicos aos patógenos invasores 4 , 5 . Os exsudados de raiz derivados de plantas, secreções, mucilagens, mucigel e lisados alteram as propriedades físico-químicas da rizosfera para atrair ou repelirMicróbios em relação aos seus anfitriões 6 . A composição química da secreção de raízes é específica da espécie, servindo assim como um filtro seletivo que permite que certos microrganismos capazes de reconhecer esses compostos florescerem na rizósfera 6 . Assim, espécies microbianas compatíveis podem ser estimuladas para ativar e melhorar suas associações, quer para benefício ou prejuízo do hospedeiro da planta 1 .
Compreender as interações planta-microbio na rizósfera é fundamental para melhorar a produtividade da planta e o funcionamento do ecossistema, uma vez que a maioria da exposição microbiana e química ocorre na estrutura radicular e na interface solo-ar 2 , 6 , 7 , 8 . No entanto, o exame das interações subterrâneas de plantas e micróbios e respostas recíprocas tem sido um desafio devido à sua intrigante Natureza complexa e dinâmica e falta de modelos experimentais adequados com estrutura radicular natural e morfologia da planta sob condições de crescimento bem controláveis. Como um dos agentes farmacêuticos mais bem estudados, Agrobacterium infecta uma grande variedade de plantas com importância agrícola e horticultural, incluindo cereja, maçã, pera, uva e rosa 9 . Agrobacterium é um organismo modelo importante para a compreensão das interações planta-patógeno e é uma ferramenta poderosa na transformação de plantas e engenharia de plantas 10 , 11 , 12 , 13 , 14 .
As interacções Molecular Plant- Agrobacterium foram bem estudadas há várias décadas e a compreensão atual da patogenicidade de Agrobacterium é extensa 9 ,F. "> 11 , 15 , 16. A patogenicidade de Agrobacterium é em grande parte atribuída às suas capacidades evoluídas de percepção de sinais derivados de plantas, resultando na modulação fina de seu programa de virulência e comunicação célula a célula, o chamado sensor de quorum 17 . O programa Agrobacterium virulence é regulado por vários sinais disponíveis na rizosfera e envolve dois conjuntos de sistemas de 2 componentes, o sistema ChvG / I eo sistema VirA / G. As condições ácidas na rizósfera ativam a transcrição de chvG / I , virA / G , E vários outros genes envolvidos na patogenicidade de Agrobacterium , incluindo virE0 , virE1 , virH1 , virH2 e genes do sistema de secreção de tipo VI (T6SS) 18. Compostos fenólicos derivados de plantas, incluindo a aceossiringona (4'-hidroxi-3 ', 5 '-dimetoxiacetofenona), ative o VSistema IrA / G de 2 componentes através de mecanismos de sinalização de fosforilação 19 . VirA / G ativa então todo o regulador vir , resultando na transferência e integração de um fragmento de DNA bacteriano de ~ 20 kb chamado DNA de transferência (DNA-T) do plasmídeo indutor de tumor (Ti) para o núcleo da planta 16 . T-DNA carrega genes responsáveis pela síntese dos hormônios vegetais ácido indol-3-acético (IAA) ( iaaM e iaaH ) e citoquinina ( ipt ), e uma vez expressa em células vegetais, produzem-se grandes quantidades dessas fitohormonas. Isso resulta em proliferação anormal de tecidos e desenvolvimento de tumores vegetais, conhecida como doença coronária, que é um problema crônico e ressurgente para as plantas 9 , 11 , 20 . IAA também atua coletivamente com ácido salicílico e ácido gamma-amino butírico para reprimir a virulência de Agrobacterium ou para reduzir Agrobacteriu M quorum sensing (QS) 17 , 21 , 22 . Para contrariar essa repressão, o T-DNA também contém genes para a biossíntese de opina, que ativa o Agrobacterium quorum detectando para promover a patogenicidade de Agrobacterium e também serve como fonte de nutrientes para o patógeno 22 , 23 .
Apesar de uma compreensão geral e profunda das interações de plantas de Agrobacterium e da transferência de T-DNA resultante para o hospedeiro da planta, os complexos eventos de sinalização na fase inicial de interação são menos bem compreendidos. Isto é parcialmente devido às limitações das abordagens convencionais para investigar a sinalização das plantas de Agrobacterium . As culturas em suspensão de células vegetais e os ferimentos artificiais específicos do local são comumente usados para estudar interações moleculares de plantas e micróbios 24 ,. ef "> 26, 27 No entanto, as suspensões de células não possuem morfologia da planta normal, em particular, as células em suspensão planta não tem estruturas de raiz e exsudatos de raízes, que são muito importantes para a activação de quimiotaxia microbiana e virulência 28, 29 A manutenção da morfologia da planta. E a estrutura da raiz foi abordada por feridas artificiais, o que facilita a infecção específica do local, resultando na detecção de genes induzidos por defesa da planta em tecido de plantas infectadas diretamente 30 , 31. Contudo, o ferimento artificial é significativamente diferente da infecção por patógenos na natureza , Particularmente como ferimento leva ao acúmulo de ácido jasmônico (JA), que interfere sistematicamente com a sinalização e defesa de plantas naturais 26. Além disso, os produtos químicos sintéticos são tipicamente usados para induzir artificialmente as respostas do hospedeiro da plantaOu virulência de patógenos. Embora seja possível a suplementação de compostos químicos que refletem as concentrações em planta , essa suplementação não explica gradualmente a difusão de exsudatos radiculares na rizósfera circundante, o que gera um gradiente quimiotático detectado pelos micróbios 28 , 32 . Dadas as limitações das abordagens convencionais para estudar as interações planta-microbio, a precisão e a profundidade dos dados obtidos podem ser impedidas e restritivas e o conhecimento gerado a partir das abordagens convencionais pode não se traduz diretamente na planta . Muitos aspectos da sinalização das plantas – Agrobacterium ainda não são totalmente compreendidos, particularmente no estágio inicial das interações, quando os sintomas da doença ainda não se desenvolveram.
Para alterar as limitações das abordagens convencionais, este trabalho apresenta um c hidroponico barato, bem controlável e flexívelSistema de ocultivação que permite aos pesquisadores obter informações mais aprofundadas sobre as vias complexas de sinalização e resposta no estágio inicial das interações de plantas-micróbios moleculares. O hidroponia tem sido amplamente utilizado para estudar nutrientes de plantas, exsudatos radiculares, condições de crescimento e os efeitos da toxicidade metálica nas plantas 33 , 34 . Existem várias vantagens dos modelos hidropônicos, incluindo os pequenos requisitos espaciais, a acessibilidade de vários tecidos vegetais, o controle rigoroso das condições nutrientes / ambientais e o controle de pragas / doenças. Os sistemas hidropônicos também são menos limitantes para o crescimento das plantas em comparação com as técnicas de revestimento em ágar / fitoagulação, que geralmente restringem o crescimento após 2-3 semanas. Importante, a manutenção de estruturas de plantas inteiras facilita a secreção de raízes naturais necessárias para a quimiotaxia microbiana e indução de virulência 8 , 29 . O sistema descriA cama aqui é mais simples e menos intensiva em mão-de-obra do que as alternativas 33 , 34 . Ele usa menos peças e não requer ferramentas além das tesouras padrão. Ele usa malha de metal (em oposição ao nylon 33 ) como um forte suporte para o crescimento da planta e um método simples de aeração sob condições estéreis através de agitação para suportar o crescimento microbiano. Além disso, o sistema pode usar malhas metálicas de vários tamanhos para suportar o crescimento da planta, que acomoda diversas espécies de plantas sem restringir a largura de suas raízes.
No sistema de cocultivação hidropônica apresentado aqui, as plantas são cultivadas em um sistema hidropônico estéril onde as raízes das plantas secretam compostos orgânicos que suportam o crescimento de bactérias inoculadas. Neste sistema de cocultivação, não são suplementados produtos químicos artificiais, como hormonas de plantas, elicitores de defesa ou produtos químicos indutores de virulência, que refletem a célula natural- assinalando a homeostase durante as interações planta-micróbio. Com este sistema de cocultivação hidropônica, foi possível determinar simultaneamente a expressão gênica no tecido da raiz de Col-0 de Arabidopsis thaliana após a infecção por Agrobacterium , bem como a ativação de genes de Agrobacterium após a cocultivação com Arabidopsis . Demonstrou-se ainda que este sistema é adequado para estudar a ligação de Agrobacterium às raízes das plantas, bem como o perfil secreto da raiz da planta, após a cocultivação (infecção) com Agrobacterium ( Figura 1 ).
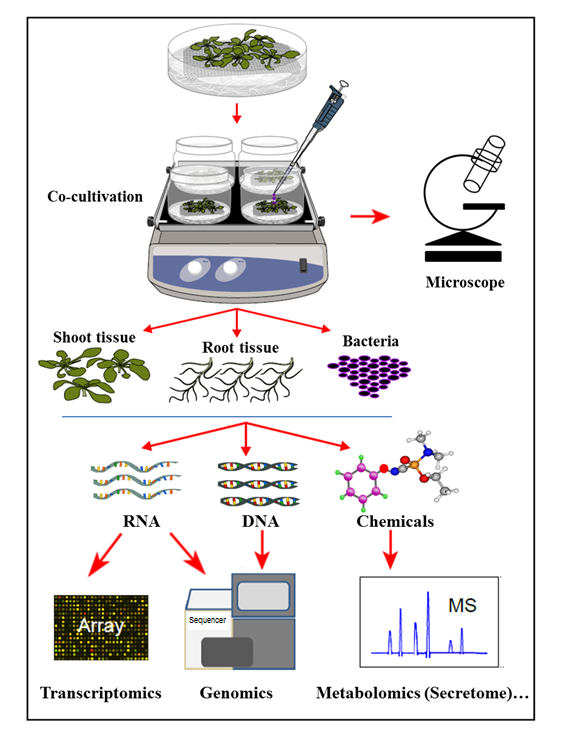
Figura 1: Visão geral do sistema de cocultivação hidropônica, com análises de amostras. As plantas são cultivadas em cima da malha (brotos acima da malha), com as raízes imersas em meio hidropônico que é então inoculado com bactérias fOu coculture. Os tecidos vegetais e as bactérias são então separados para extrações e análises simultâneas. Esta figura foi modificada a partir da referência 35 .
