Los microbios asociados a las plantas desempeñan un papel importante en el ciclo biogeoquímico, la biorremediación, la mitigación del cambio climático, el crecimiento y la salud de las plantas y la tolerancia de las plantas a las tensiones bióticas y abióticas. Los microorganismos interactúan con las plantas tanto directamente a través del contacto de la pared celular de la planta como indirectamente a través de la secreción y señalización química 1 , 2 , 3 . Como organismos sésiles, las plantas han desarrollado mecanismos directos e indirectos para resistir la infección por patógenos. Las defensas directas incluyen defensas estructurales y la expresión de proteínas de defensa, mientras que las defensas indirectas incluyen la producción secundaria de metabolitos de plantas y la atracción de organismos antagónicos a patógenos invasores 4 , 5 . Los exudados radiculares, secreciones, mucílagos, mucigel y lisados de las raíces alteran las propiedades físico-químicas de la rizosfera para atraer o repelerMicrobios hacia sus huéspedes 6 . La composición química de la secreción de las raíces es específica de la especie, sirviendo así como un filtro selectivo que permite que ciertos microorganismos capaces de reconocer tales compuestos florezcan en la rizosfera 6 . Por lo tanto, las especies microbianas compatibles pueden ser estimuladas para activar y mejorar sus asociaciones, ya sea en beneficio o detrimento de la planta huésped 1 .
La comprensión de las interacciones planta-microbio en la rizosfera es clave para mejorar la productividad de la planta y el funcionamiento del ecosistema, ya que la mayoría de la exposición microbiana y química ocurre en la estructura de la raíz y en la interfase suelo-aire 2 , 6 , 7 , 8 . Sin embargo, el examen de las interacciones planta-microbios subterráneas y respuestas recíprocas ha sido un desafío debido a su intrigante Compleja y dinámica y la falta de modelos experimentales adecuados con estructura de raíces naturales y morfología de plantas bajo condiciones de crecimiento controlables. Como uno de los fitopatógenos más estudiados, Agrobacterium infecta una amplia gama de plantas con importancia agrícola y hortícola, incluyendo cereza, manzana, pera, uva y rosa 9 . Agrobacterium es un organismo modelo importante para la comprensión de las interacciones planta-patógeno y es una herramienta poderosa en la transformación de plantas y la ingeniería de plantas 10 , 11 , 12 , 13 , 14 .
Las interacciones moleculares entre plantas y Agrobacterium han sido bien estudiadas durante varias décadas, y la comprensión actual de la patogenicidad de Agrobacterium es extensa 9 ,f "> 11, 15, 16. patogenicidad Agrobacterium se atribuye en gran parte a sus capacidades evolucionadas de percibir señales derivadas de plantas, dando como resultado la modulación fina de su programa de la virulencia y la comunicación de célula a célula, llamado quórum de detección 17. La El programa de virulencia de Agrobacterium está regulado por varias señales disponibles en la rizosfera e incluye dos sistemas de sistemas de dos componentes, el sistema ChvG / I y el sistema VirA / G. Las condiciones ácidas en la rizosfera activan la transcripción de chvG / I , virA / G , Y varios otros genes implicados en la patogenicidad de Agrobacterium , incluyendo virE0 , virE1 , virH1 , virH2 y genes del sistema de secreción de tipo VI (T6SS) 18. Compuestos fenólicos derivados de plantas, incluyendo acetosiringona (4'-hidroxi-3 ', 5 '-dimetoxiacetofenona), activar el VIrA / G sistema de 2 componentes a través de mecanismos de señalización de fosforilación 19 . VirA / G activa entonces todo el vir regulon, dando como resultado la transferencia e integración de un fragmento de ADN bacteriano de aproximadamente 20 kb denominado ADN de transferencia (T-DNA) desde su plásmido inductor de tumores (Ti) al núcleo de la planta 16 . T-DNA transporta genes responsables de la síntesis de las hormonas vegetales indol-3-acético (IAA) ( iaaM y iaaH ) y citoquinina ( ipt ), y una vez expresado en las células vegetales, se producen grandes cantidades de estas fitohormonas. Esto da lugar a la proliferación anormal del tejido y al desarrollo del tumor de la planta, conocida como enfermedad de la bilis de la corona, que es un problema crónico y resurgente para las plantas 9 , 11 , 20 . IAA también actúa colectivamente con ácido salicílico y ácido gamma-amino butírico para reprimir la virulencia de Agrobacterium o para reducir Agrobacteriu M quorum sensing (QS) 17 , 21 , 22 . Para contrarrestar esta represión, el T-DNA también lleva genes para la biosíntesis de opina, que activa la detección de Agrobacterium quorum para promover la patogenicidad de Agrobacterium y también sirve como fuente de nutrientes para el patógeno 22 , 23 .
A pesar de una profunda comprensión global de Agrobacterium- planta interacciones y la transferencia de T-DNA resultante en el huésped de la planta, los complejos eventos de señalización en la etapa inicial de la interacción son menos bien entendidos. Esto se debe en parte a las limitaciones de los métodos convencionales para investigar la señalización de planta de Agrobacterium . Los cultivos de suspensión de células vegetales y heridas artificiales específicas del sitio se usan comúnmente para estudiar las interacciones moleculares planta-microbio 24 ,. ef "> 26, 27 Sin embargo, las suspensiones de células carecen de morfología de la planta típica, en particular, células en suspensión planta no tienen estructuras profundas y los exudados de la raíz, que son muy importantes para la activación de la quimiotaxis microbiana y la virulencia 28, 29 El mantenimiento de la morfología de la planta. Y la estructura de la raíz se ha tratado de herir artificialmente a las plantas, lo que facilita la infección específica del sitio, lo que resulta en la detección de genes inducidos relacionados con la defensa de la planta en tejido vegetal directamente infectado 30 , 31. Sin embargo, la herida artificial es significativamente diferente de la infección patógena en la naturaleza , Particularmente como herida conduce a la acumulación de ácido jasmónico (JA), que sistemáticamente interfiere con la señalización de la planta natural y la defensa 26. Además, los productos químicos sintéticos se utilizan normalmente para inducir artificialmente las respuestas del huésped de la plantaO virulencia patógena. Aunque la suplementación de tales compuestos químicos refleja concentraciones en planta es posible, tal suplementación no explica la difusión de los exudados de raíz gradualmente en la rizosfera circundante, lo que genera un gradiente quimiotáctico detectado por los microbios 28 , 32 . Dadas las limitaciones de los enfoques convencionales para estudiar las interacciones planta-microbios, la exactitud y la profundidad de los datos obtenidos podrían ser impedidas y restrictivas, y el conocimiento generado a partir de los enfoques convencionales puede no traducir directamente in planta . Muchos aspectos de la señalización de Agrobacterium en las plantas aún no se entienden completamente, particularmente en la etapa temprana de las interacciones, cuando los síntomas de la enfermedad aún no se han desarrollado.
Para enmendar las limitaciones de los enfoques convencionales, este trabajo presenta un bajo costo, controlable, y flexible hydroponic cOcultivation sistema que permite a los investigadores a obtener una visión más profunda de la compleja de señalización y respuesta a las vías en la etapa inicial de la planta molecular microbio interacciones. La hidroponía se ha utilizado ampliamente para estudiar los nutrientes de las plantas, los exudados de las raíces, las condiciones de crecimiento y los efectos de la toxicidad metálica en las plantas 33 , 34 . Existen varias ventajas de los modelos hidropónicos, incluyendo las pequeñas exigencias espaciales, la accesibilidad de los diversos tejidos vegetales, el estricto control de las condiciones nutrientes / ambientales y el control de plagas / enfermedades. Los sistemas hidropónicos también son menos limitantes para el crecimiento de las plantas en comparación con las técnicas de placas de agar / phytoagar, que típicamente restringen el crecimiento después de 2-3 semanas. Es importante destacar que el mantenimiento de las estructuras de toda la planta facilita la secreción de raíces naturales necesarias para la quimiotaxis microbiana y la inducción de la virulencia [ 8 , 29] . El sistema descriLa cama aquí es más simple y menos laboriosa que las alternativas 33 , 34 . Utiliza menos piezas y no requiere ninguna otra herramienta que las tijeras estándar. Utiliza malla metálica (en oposición al nylon 33 ) como soporte fuerte para el crecimiento de la planta y un método simple de aireación en condiciones estériles mediante agitación para soportar el crecimiento microbiano. Además, el sistema puede utilizar malla metálica de varios tamaños para apoyar el crecimiento de la planta, que acomoda diversas especies de plantas sin restringir la anchura de sus raíces.
En el sistema de cocultivación hidropónica presentado aquí, las plantas se cultivan en un sistema hidropónico estéril donde las raíces de las plantas secretan compuestos orgánicos que apoyan el crecimiento de las bacterias inoculadas. En este sistema de cocultivación, no se suplementan productos químicos artificiales, tales como hormonas vegetales, elicitor de defensa, o productos químicos que inducen virulencia, lo que refleja la célula natural-significando la homeostasis durante las interacciones planta-microbio. Con este sistema de cocultivación hidropónica, fue posible determinar simultáneamente la expresión génica en el tejido de raíz de Col-0 de Arabidopsis thaliana tras la infección por Agrobacterium , así como la activación de genes de Agrobacterium tras la cocultivación con Arabidopsis . Se demostró además que este sistema es adecuado para estudiar la fijación de Agrobacterium a las raíces de las plantas, así como el perfil de secreto de la raíz de la planta, al cocultivar (infección) con Agrobacterium ( Figura 1 ).
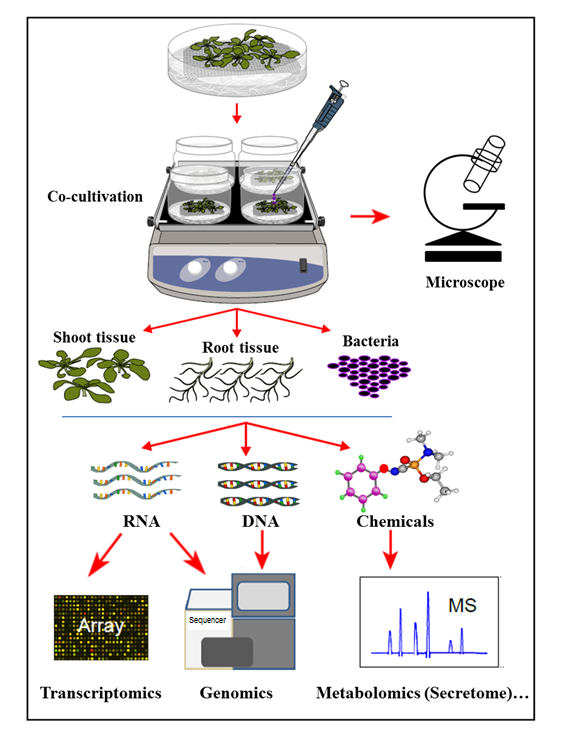
Figura 1: Visión general del sistema de cocultivación hidropónica, con análisis de muestras. Las plantas se cultivan en la parte superior de la malla (brotes por encima de la malla), con las raíces sumergidas en medio hidropónico que luego se inocula con bacterias fO el coculture. Los tejidos vegetales y las bacterias se separan para extracciones y análisis simultáneos. Esta cifra ha sido modificada a partir de la referencia 35 .
