Bitki ile ilişkili mikrop biyojeokimyasal döngü, biyolojik giderme, iklim değişikliğinin hafifletilmesi, bitki büyümesi ve sağlığı ve biyotik ve abiyotik streslere karşı bitki toleransında önemli rol oynamaktadır. Mikroorganizmalar bitkilerle doğrudan bitki hücresi duvar teması yoluyla ve dolaylı olarak kimyasal salınım ve sinyalleme 1 , 2 , 3 aracılığıyla etkileşirler. Sabit organizmalar olarak, bitkiler patojenler tarafından enfeksiyona karşı doğrudan ve dolaylı mekanizmalar geliştirmiştir. Dolaylı savunma ikincil bitki metabolit üretimini ve patojenleri 4, 5 istila antagonistik organizmaların gözde içerisinde ise direkt savunma yapısal savunma ve savunma proteinlerinin ekspresyonunu içerir. Bitki kökenli kök sızıntıları, salgıları, müsilatları, müsigel ve lizatları rizosferin fiziksel-kimyasal özelliklerini çekme veya itme için değiştirirEv sahiplerine karşı mikrop 6 . Kök sekresyonunun kimyasal bileşimi, türe özgüdür, böylelikle bu bileşiklerin tanınması için yeterli mikroorganizmaların rizosfer 6'da gelişmesine izin veren seçici bir filtre görevini görür. Böylece uyumlu bir mikrobik türler de, bitki konakçıya 1 yararına veya zararına, aktif ve ilişkilerini geliştirmek için uyarılabilir.
Mikrobiyal ve kimyasal maruziyetin büyük bir kısmı kök yapısı ve toprak-hava arayüzü 2 , 6 , 7 , 8'de gerçekleştiği için, rizosferdeki bitki-mikrop etkileşimlerini anlamak, bitki verimliliğini ve ekosistemi işlerliği güçleştirmenin anahtarıdır. Bununla birlikte, yeraltı bitki-mikrop etkileşimlerinin ve karşılıklı tepkilerin incelenmesi, ilginç bir şekilde Karmaşık ve dinamik doğası ve sıkı kontrol edilebilir büyüme koşulları altında doğal kök yapısı ve bitki morfolojisi ile uygun deneysel modellerin olmaması. En ağır çalışılan fitopatojenlere biri olan Agrobacterium kiraz, elma, armut, üzüm dahil tarım ve bahçecilik öneme sahip bitkilerin geniş bir yelpazede bozar ve 9 yükseldi. Agrobacterium anlama bitki-patojen etkileşimlerinin önemli bir model organizma ve bitki transformasyonu ve tesis mühendisliği 10, 11, 12, 13, 14 bir güçlü bir araçtır.
Moleküler bitki- Agrobacterium etkileşimleri birkaç on yıl boyunca iyi çalışılmış ve Agrobacterium patojenisitesinin mevcut anlayışı geniş kapsamlı 9 ,F "> 11 , 15 , 16. Agrobacterium patojenitesi, büyük oranda, bitki kaynaklı sinyalleri algılama yetenekleri ve bunun virulans programının ve hücre-hücre iletişiminin ince modülasyonuyla sonuçlanan, yeterlilik nütrisyonu 17 olarak adlandırılan şekilde ortaya çıktığı anlamına gelmektedir. Agrobacterium virulence programı, rizosferdeki birkaç sinyal ile düzenlenir ve iki bileşenli sistemlerden oluşan ChvG / I sistemi ve VirA / G sistemini içerir: Rizosferdeki asidik koşullar, chvG / I , virA / G'nin transkripsiyonunu aktive eder ve virE0, virE1, virH1, virH2 ve tip VI salgılama sistemine (T6SS) 18. genleri de dahil olmak üzere, Agrobacterium patojenik, kapsanan çeşitli diğer genler asetosiringon (4'-hidroksi-3' , 5 de dahil olmak üzere fenolik bileşikleri, bitki-türevi '-dimetoksiasetofenon), V'yi aktive edinIrA / G 2 komponentli sistem fosforilasyon sinyal mekanizmaları vasıtasıyla 19 . VirA / G daha sonra tüm vir regülatörünü aktive eder ve transfer DNA'sı (T-DNA) olarak adlandırılan ~ 20 kb bakteri DNA fragmanının tümör kaynaklı (Ti) plasmidinden bitki çekirdeğine 16 aktarılması ve entegrasyonu ile sonuçlanır. T-DNA, indol-3-asetik asit (IAA) ( iaaM ve iaaH ) ve sitokinin ( ipt ) bitki hormonlarının sentezinden sorumlu genleri taşır ve bir zamanlar bitki hücrelerinde eksprese edildiğinde, bu bitkisel hormonların büyük miktarları üretilir. Bu, anormal doku çoğalması ve bitki tümörü gelişimine, yani 9 , 11 , 20 nolu bitkiler için kronik ve yeniden dirilme problemi olan kron kalp hastalığı ile sonuçlanır. IAA da Agrobacterium virulence'ı baskılamak veya Agrobacteriu'yu azaltmak için salisilik asit ve gama-amino bütirik asit ile birlikte hareket eder M zayıflama algılama (QS) 17 , 21 , 22 . Bu baskıya karşı koymak için, T-DNA da Agrobacterium patojenisitesini teşvik etmek için Agrobacterium çekirdeğini aktive eden ve ayrıca patojen 22 , 23 için bir besin kaynağı olarak görev yapan opine biyosentezi genleri taşır.
Agrobacterium -plant etkileşimleri ve bitki konakçığına dönüşen T-DNA transferinin derinlemesine anlaşılmasına rağmen, etkileşimin ilk aşamasındaki kompleks sinyal olayları daha az anlaşılır. Bu, kısmen Agrobacterium -plant sinyalizasyonunu araştıran konvansiyonel yaklaşımların kısıtlamalarına bağlı. Bitki hücre süspansiyon kültürleri ve suni bölgeye özgü yaralama, moleküler bitki-mikrop etkileşimleri 24 ,Ef "> 26 , 27. Ancak, hücre süspansiyonları tipik bitki morfolojisine sahip değildir, özellikle bitki süspansiyon hücreleri kök yapısı ve kök sızıntısı içermez ve bunlar mikrobiyal kemotaksis ve virülence aktive etmek için çok önemlidir 28 , 29. Bitki morfolojisinin korunması Ve kök yapısı, doğrudan enfekte bitki dokusunda 30 , 31 endüklenen bitki savunma ile ilgili genlerin saptanmasına neden olarak, bölgeye özgü enfeksiyonu kolaylaştıran yapay olarak yaralamış bitkiler tarafından ele alınmıştır.Ancak suni yaralanma, doğadaki patojen enfeksiyonundan belirgin olarak farklıdır , Çünkü özellikle yaralanma, doğal bitki sinyalizasyonunu ve savunmasını sistemik olarak engelleyen jasmonik asit (JA) birikimine neden olarak 26. Ayrıca, sentetik kimyasal maddeler tipik olarak bitki konukçu yanıtlarını yapay olarak indüklemek için kullanılırVeya patojen virulence. Bu tür kimyasal bileşiklerin planta konsantrasyonları yansıtacak şekilde takviye edilmesi mümkün olsa da, bu takviye, kök sızıntılarının, mikroskoplar 28 , 32 tarafından algılanan bir kemotaktik gradyan oluşturan çevresindeki rizosfer içerisindeki kademeli olarak difüzyonunu hesaba katmaz. Bitki-mikrop etkileşimlerinin incelenmesine yönelik geleneksel yaklaşımların sınırlamaları göz önüne alındığında, elde edilen verilerin doğruluğu ve derinliği engellenebilir ve kısıtlayıcı olabilir ve konvansiyonel yaklaşımlardan üretilen bilgiler doğrudan planta tercüme edilemez. Bitki- Agrobacterium sinyalizasyonunun bir çok yönü, özellikle hastalık semptomları henüz gelişmediğinde, etkileşimlerin erken evresinde tam olarak anlaşılamamıştır.
Konvansiyonel yaklaşımların sınırlamalarını değiştirmek için, bu çalışma, ucuz, sıkı kontrol edilebilir ve esnek hidroponik cAraştırmacılar moleküler bitki-mikrop etkileşimlerinin ilk aşamasında karmaşık sinyalizasyon ve yanıt yollarına daha derin kavrayışlar kazandıran bir ocultivation sistemi. Hidroponik bitki besin maddeleri, kök sızıntıları, büyüme koşulları ve metal toksisitesinin bitkiler üzerindeki etkilerini incelemek için yaygın bir şekilde kullanılmaktadır 33 , 34 . Küçük mekan gereksinimleri, çeşitli bitki dokularının erişilebilirliği, besin / çevre koşullarının sıkı kontrolü ve zararlı / hastalık kontrolü dahil olmak üzere hidrofobik modellerin pek çok avantajı vardır. Ayrıca, 2-3 hafta sonra büyümeyi sınırlayan agar / fitoagar kaplama teknikleriyle karşılaştırıldığında, hidrofobik sistemler bitki büyümesini daha az sınırlandırmaktadır. Önemli olarak, bütün bitki yapılarının bakım mikrobiyal kemotaksisi ve hastalık oluşturma indüksiyonu 8, 29 için gerekli doğal kök sekresyonunu kolaylaştıran. Sistem açıklamasıBurada yatak 33 ve 34 seçeneklerinden daha basit ve daha az emek-yoğun. Daha az parça kullanır ve standart makas haricinde herhangi bir alet gerektirmez. Bitki büyümesi için güçlü bir destek ve mikrobik büyümeyi desteklemek için sallayarak steril koşullar altında basit bir havalandırma yöntemi olarak metal örgü kullanır (naylon 33'e karşı). Buna ek olarak, sistem, köklerinin genişliğini sınırlamaksızın çeşitli bitki türlerini barındıran bitki büyümesini desteklemek için çeşitli ebatlarda metal örgü kullanabilir.
Burada sunulan hidroponik kokültivasyon sisteminde bitkiler, bitki köklerinin aşılanmış bakterilerin büyümesini destekleyen organik bileşikleri salgılayan steril bir hidroponik sistemde yetiştirilir. Bu ko-yetiştirme sisteminde, bitki hormonları, savunma elitörü veya virulans indükleyici kimyasallar gibi yapay kimyasallar eklenmemektedir; bu da doğal hücrenin-Bitki-mikrop etkileşimleri sırasında homeostazı işaretleme. Bu hidroponik kokültivasyon sistemi ile, Agrobacterium tarafından enfeksiyon üzerine Arabidopsis thaliana Col-0 kökü dokusunda gen ifadesinin yanı sıra Arabidopsis ile birlikte yetiştirme üzerine Agrobacterium genlerinin aktivasyonu aynı anda belirlenebilirdi. Bundan başka, bu sistem, Agrobacterium (Şekil 1) ile birlikte kültürün (enfeksiyon) üzerine, kökler, aynı zamanda, bitki kök secretome profilini bitki Agrobacterium eki incelemek için uygun olduğu gösterilmiştir.
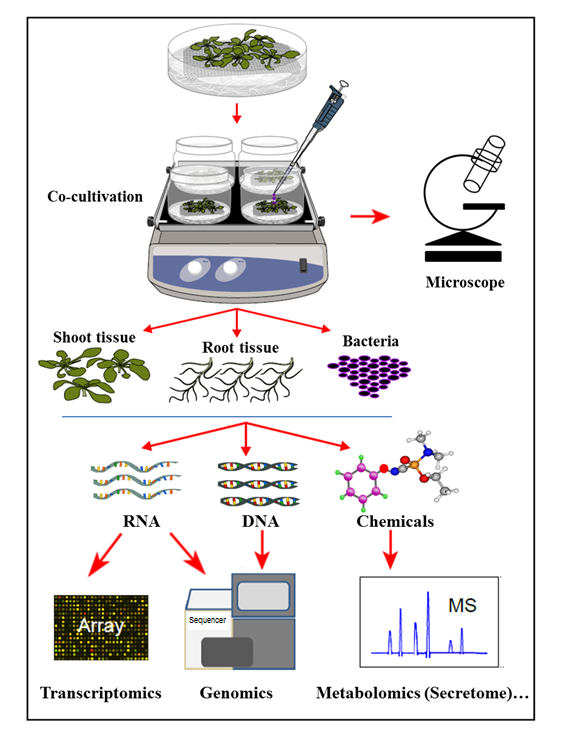
Şekil 1: Örnek Analizlerle Hidroponik Kokültürasyon Sistemine Genel Bakış. Bitkiler, ağın üstünde yetiştirilir (ağın üstünde sürgünler), kökleri hidrofobik ortamda daldırıp daha sonra bakteri fVeya kokültür. Bitki dokuları ve bakteriler daha sonra eşzamanlı ekstraksiyon ve analizler için ayrılır. Bu rakam referans 35'den değiştirildi.
