Les microbes associés aux plantes jouent un rôle important dans le cyclisme biogéochimique, la bioremédiation, l'atténuation des changements climatiques, la croissance et la santé des plantes et la tolérance des plantes aux contraintes biotiques et abiotiques. Les microorganismes interagissent avec les plantes directement à travers le contact de la paroi cellulaire de la plante et indirectement par la sécrétion chimique et la signalisation 1 , 2 , 3 . En tant qu'organes sessiles, les plantes ont développé des mécanismes directs et indirects pour résister à l'infection par des agents pathogènes. Les défenses directes comprennent les défenses structurelles et l'expression des protéines de défense, tandis que les défenses indirectes incluent la production secondaire de métabolites de plantes et l'attraction d'organismes antagonistes aux pathogènes envahissants 4 , 5 . Les exsudats de racines dérivés des plantes, les sécrétions, les mucilages, les mucigel et les lysats modifient les propriétés physico-chimiques de la rhizosphère pour attirer ou repousserDes microbes vers leurs hôtes 6 . La composition chimique de la sécrétion des racines est spécifique à l'espèce, servant ainsi de filtre sélectif qui permet à certains microorganismes capables de reconnaître de tels composés se développer dans la rhizosphère 6 . Ainsi, des espèces microbiennes compatibles peuvent être stimulées pour activer et améliorer leurs associations, soit au bénéfice ou au détriment de l'hôte 1 de la plante.
La compréhension des interactions plantes-microbes dans la rhizosphère est essentielle pour améliorer la productivité des plantes et le fonctionnement de l'écosystème, car une majorité de l'exposition microbienne et chimique se produit à la structure racine et à l'interface sol-air 2 , 6 , 7 , 8 . Cependant, l'examen des interactions souterraines des plantes-microbes et des réponses réciproques a été un défi en raison de son intrigué La nature complexe et dynamique et le manque de modèles expérimentaux appropriés avec structure racine naturelle et morphologie végétale dans des conditions de croissance très contrôlables. Comme l'un des phytopathogènes les plus fortement étudiés, Agrobacterium infecte un large éventail de plantes d'importance agricole et horticole, y compris la cerise, la pomme, la poire, le raisin et la rose 9 . Agrobacterium est un organisme modèle important pour la compréhension des interactions plantes-pathogènes et est un outil puissant dans la transformation des plantes et l'ingénierie des usines 10 , 11 , 12 , 13 , 14 .
Les interactions Molecular Plant- Agrobacterium ont été bien étudiées depuis plusieurs décennies et la compréhension actuelle de la pathogenèse d' Agrobacterium est importante 9 ,F "> 11 , 15 , 16. La pathogénicité de l' agrobacterium est largement attribuée à ses capacités évoluées de perception des signaux dérivés des plantes, ce qui entraîne la modulation fine de son programme de virulence et la communication cellulaire à cellulaire, appelée détection de quorum 17 . Le programme Agrobacterium virulence est réglementé par plusieurs signaux disponibles dans la rhizosphère et implique deux ensembles de systèmes à 2 composants, le système ChvG / I et le système VirA / G. Les conditions acides de la rhizosphère activent la transcription de chvG / I , virA / G , Et plusieurs autres gènes impliqués dans la pathogénicité d' Agrobacterium , y compris virE0 , virE1 , virH1 , virH2 et les gènes du système de sécrétion de type VI (T6SS) 18. Composés phénoliques dérivés des plantes, y compris l'acétosyringone (4'-hydroxy-3 ', 5 '-diméthoxyacétophénone), activez le VSystème à 2 composants irA / G par des mécanismes de signalisation de phosphorylation 19 . VirA / G active alors tout le régulateur vir , ce qui entraîne le transfert et l'intégration d'un fragment d'ADN bactérien de ~ 20 kb appelé ADN de transfert (ADN-T) à partir de son plasmide inducteur de tumeur (Ti) dans le noyau de la plante 16 . T-DNA porte des gènes responsables de la synthèse des hormones végétales de l'acide indole-3-acétique (IAA) ( iaaM et iaaH ) et de la cytokinine ( ipt ), et une fois exprimée dans les cellules végétales, de grandes quantités de ces phytohormones sont produites. Il en résulte une prolifération anormale des tissus et un développement de la tumeur végétale, connue sous le nom de maladie de la couronne, qui est un problème chronique et résurgent pour les plantes 9 , 11 , 20 . IAA agit également collectivement avec de l'acide salicylique et de l'acide gamma-amino butyrique pour réprimer Agrobacterium virulence ou pour réduire Agrobacteriu M quorum sensing (QS) 17 , 21 , 22 . Pour contrer cette répression, l'ADN-T porte également des gènes pour la biosynthèse de l'opine, qui active Agrobacterium quorum qui détecte la pathogénicité d' Agrobacterium et sert également de source nutritive pour le pathogène 22 , 23 .
En dépit d'une compréhension générale globale des interactions avec les plantes d' Agrobacterium et du transfert d'ADN-T résultant dans l'hôte de la plante, les événements de signalisation complexes au stade initial de l'interaction sont moins bien compris. Ceci est en partie dû aux limites des approches classiques pour enquêter sur la signalisation des plantes Agrobacterium . Les cultures en suspension de cellules végétales et les blessures artificielles spécifiques au site sont couramment utilisées pour étudier les interactions moléculaires-microbes 24 ,. ef "> 26, 27 Cependant, les suspensions cellulaires manquent la morphologie des plantes typiques, en particulier, les cellules de suspension des plantes ne sont pas des structures profondes et exsudats racinaires, qui sont très importants pour l' activation chimiotactisme microbienne et la virulence 28, 29 Le maintien de la morphologie des plantes. Et la structure racine a été traitée par des plantes blessant artificiellement, ce qui facilite l'infection spécifique au site, ce qui entraîne la détection de gènes induits par la défense des plantes dans les tissus végétaux infectés directement 30 , 31. Cependant, les blessures artificielles sont significativement différentes de l'infection pathogène dans la nature , D'autant plus que les blessures entraînent une accumulation d'acide jasmonique (JA), qui interfère systémiquement avec la signalisation et la défense des plantes naturelles 26. En outre, les produits chimiques de synthèse sont généralement utilisés pour induire artificiellement les réponses de l'hôte de la planteOu la virulence des agents pathogènes. Bien que la supplémentation de tels composés chimiques reflétant les concentrations en planta soit possible, une telle supplémentation ne tient pas compte de la diffusion des exsudats radiculaires progressivement dans la rhizosphère environnante, ce qui génère un gradient chimiotactique détecté par les microbes 28 , 32 . Compte tenu des limites des approches conventionnelles pour étudier les interactions entre les plantes et les microbes, la précision et la profondeur des données obtenues pourraient être entravées et restrictives, et les connaissances générées par les approches conventionnelles ne peuvent pas se traduire directement en plan . De nombreux aspects de la signalisation des plantes- Agrobacterium ne sont pas encore entièrement compris, en particulier au stade précoce des interactions, lorsque les symptômes de la maladie ne sont pas encore développés.
Pour modifier les limites des approches conventionnelles, ce travail présente un hydroponique c caisse peu coûteux, étroitement contrôlable et flexibleSystème d'ocultivation qui permet aux chercheurs d'acquérir des connaissances plus approfondies sur les voies complexes de signalisation et de réponse au stade initial des interactions moléculaire-microbe moléculaire. La culture hydroponique a été largement utilisée pour étudier les nutriments des plantes, les exsudats radiculaires, les conditions de croissance et les effets de la toxicité métallique sur les plantes 33 , 34 . Il existe plusieurs avantages des modèles hydroponiques, y compris les petites exigences spatiales, l'accessibilité de divers tissus végétaux, le contrôle strict des conditions nutritives / environnementales et le contrôle des ravageurs / maladies. Les systèmes hydroponiques sont également moins limitatifs pour la croissance des plantes par rapport aux techniques de gélose / phytoagar, ce qui limite généralement la croissance après 2-3 semaines. Fait important, le maintien de structures entières facilite la sécrétion naturelle des racines nécessaire à la chimiotaxie microbienne et à l'induction de la virulence 8 , 29 . Le système décriLe lit ici est plus simple et moins exigeant en main-d'œuvre que les alternatives 33 , 34 . Il utilise moins de pièces et ne nécessite aucun outil autre que les ciseaux standard. Il utilise des mailles métalliques (par opposition au nylon 33 ) comme un solide support pour la croissance des plantes et une méthode simple d'aération dans des conditions stériles en secouant pour soutenir la croissance microbienne. En outre, le système peut utiliser des grilles métalliques de différentes tailles pour soutenir la croissance des plantes, ce qui permet d'inclure diverses espèces de plantes sans limiter la largeur de leurs racines.
Dans le système de cocultivation hydroponique présentée ici, les plantes sont cultivées dans un système hydroponique stérile où les racines des plantes sécrètent des composés organiques soutenant la croissance des bactéries inoculées. Dans ce système de cocultivation, aucun produit chimique artificiel, tel que des hormones végétales, un élicateur de défense ou des produits chimiques inductifs de la virulence, est complété, ce qui reflète la cellule naturelle- signification de l'homéostasie lors des interactions plantes-microbes. Avec ce système de cocultivation hydroponique, il a été possible de déterminer simultanément l'expression des gènes dans le tissu racidique d'Arabidopsis thaliana Col-0 lors d'une infection par Agrobacterium , ainsi que l'activation des gènes Agrobacterium lors de la cocultivation avec Arabidopsis . Il a également été démontré que ce système est approprié pour étudier l'attachement d' Agrobacterium aux racines des plantes, ainsi que le profil de la racine de la plante, lors de la cocultivation (infection) avec Agrobacterium ( Figure 1 ).
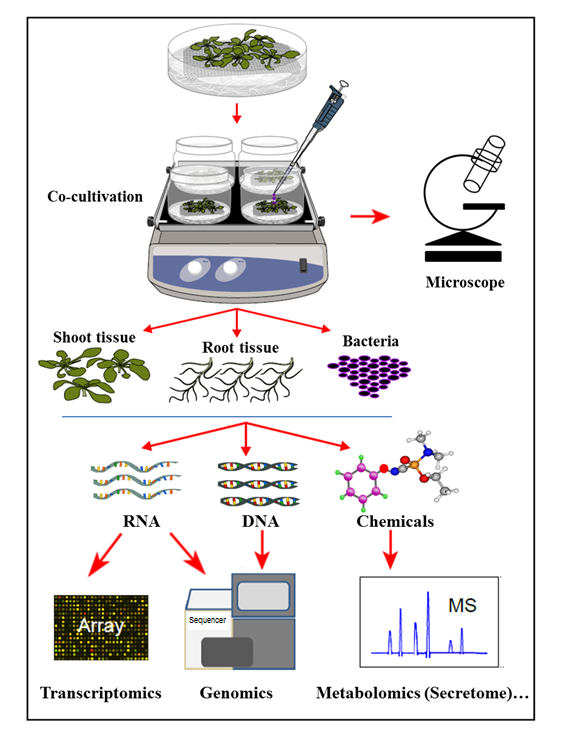
Figure 1: Vue d'ensemble du système de cocultivation hydroponique, avec des analyses d'échantillons. Les plantes sont cultivées au-dessus du maillage (pousses au-dessus du maillage), les racines étant immergées dans un milieu hydroponique qui est ensuite inoculé avec des bactéries fOu coculture. Les tissus végétaux et les bactéries sont ensuite séparés pour des extractions et des analyses simultanées. Ce chiffre a été modifié à partir de la référence 35 .
