Hipóxia, ou seja, dissolvido as concentrações de oxigênio [] suficientemente baixo para afetar negativamente biológicas e processos ecológicos, mas muitas vezes funcionalmente definida como [fazer] < 2 mg / L1e anóxia (funcionalmente definida como [fazer] de 0.0-0.2 mg/L) ocorrem mais frequentemente e severamente em águas costeiras do mundo, estuários e o oceano profundo2,3 e são muitas vezes agravada pela crescente eutrofização4,5. Com uma extensão de areal crescente de hipóxia e anóxia, macrofauna são afetados negativamente e perder a medida do habitat e qualidade de habitat. Mudança climática prevê-se a agravar a hipóxia e anóxia6.
Em muitos estuários estratificados, enriquecidos com nutrientes como Chesapeake Bay, Estados Unidos, hipóxia sazonalmente persistente pode prevalecer e pode ocorrer ano após ano2. Além disso, diel-ciclismo de hipóxia é frequente em estuários como a Baía de Chesapeake e outros locais e ocorre tarde durante a noite ou de madrugada no verão7,8.
A maioria dos estudos centraram-se sobre os efeitos da exposição contínua dos organismos para baixo [fazer] e sua tolerância à hipóxia e anóxia9,10,11,12,13,14 . Além disso, estudos têm olhou para a mudança em grande escala na composição de espécies distribuições, abundâncias e espécies em resposta ao prolongado baixa [fazer]4,15. Frequentemente espécies que são muito sensíveis à baixa [,] morrer em massas,16 deslocando as restantes espécies de uma fauna mais jovem, de menor porte, curta duração, como, por exemplo, encontradas no ecossistema Louisiana-Texas prateleira4.
Mudanças comportamentais normalmente precedem comunidade colapso17 e estudos relataram sobre respostas comportamentais dos organismos para estendido baixa [fazer]4,16,17,18,19 ,20,21,22,23,24,25. Estes estudos, no entanto, não se concentre as respostas dos organismos para exposições diel-ciclismo de hipóxia e a natureza flutuante do [fazer] disponibilidade em estuários.
Hipoxia Diel-ciclismo em estuários rasos tem recebido crescente sensibilização como estudos monitorar [fazer] mais frequentemente ao longo dos dias com sondas em estuários16,26. Água pode permanecer hipóxica por horas no final da noite ou de madrugada no verão quando não há nenhuma fotossíntese geradora de oxigênio durante a noite mas o alto consumo de oxigênio de7,de respiração aeróbia16. Também foi encontrado que as marés afetou o diel ciclismo de baixas condições de fazer com a mais extremas mínimos observados quando as marés baixa coincidiu com o fim da noite27. Só depois de várias horas de hipóxia [fazer] voltar a normoxia7,16,28 do ciclo diário.
Para determinar a resposta comportamental de c. virginica hipóxia diel-ciclismo e pH nós monitorados a abertura e fechamento das válvulas de ostras expostas ao laboratório induzido diel cíclico e ciclismo de [fazer] pH. Gape respostas de bivalves têm sido utilizadas para detectar condições ambientais adversas. Tampas de válvula de bivalves em resposta a contaminantes29,30,31, algas tóxicas32,33,34, poluição térmica35,36 , 37,38,39,40, alimentação regime39,41, emersão37,42, fotoperíodo43 diminuir a quantidade de alimentos , 44, pH45,46e combinado de pH e oxigênio dissolvido47 foram medidos. Técnicas de abertura, por exemplo, incluíram observações diretas48,49,13, medições contínuas usando ímãs (Dreissena monitor)50e reed switches ou sensores de fibra óptica 51 que requerem água clara. Além disso, sensores de Hall de força magnética e campo magnético têm sido usados para estudar o mexilhão gape ângulo52,53,54,55e um sistema de indução eletromagnética de alta frequência que pode, a medida da variação da distância entre duas bobinas elétricas que são coladas nas válvulas foi usado56,57,58,59. Uma fonte de alta tensão é necessária para o sistema de indução eletromagnética e poder tem que ser entregue para ambos os lados do escudo52. Este sistema também é comercialmente disponível como o “MOSSELMONITOR” (http://mosselmonitor.nl/).
Em um orçamento apertado pesquisa, construímos um monitor barato Extensômetro (SGM) para medir continuamente gape ostra sobre induzida em laboratório diel ciclismo de [fazer] e pH, em condições de baixa visibilidade. Nosso sistema também é muito mais simples do que os sistemas concorrentes, permitindo que muitos animais para serem instrumentados durante um experimento. Nós queríamos determinar as respostas comportamentais de c. virginica para diel ciclismo severa ([] = 0,6 mg/L) hipóxia com pH de controle (pH = 7,8) e ciclagem de pH (pH = 7,8-7.0), respectivamente e gape respostas a suave ([] = 1,7 mg/L) hipóxia. Além disso, queríamos determinar se as ostras são capazes de responder rapidamente às mudanças em [] sobre o diel ciclo e como eles respondem quando normoxia retorna após um evento hipóxico. Talvez as ostras são otimamente adaptadas ao ambiente rapidamente flutuante que é encontrado em muitos estuários16,27 onde eles vivem. Enquanto mais complexa válvula gape monitores estão disponíveis, a SGM oferece uma técnica de baixo custo que permite medições contínuas de válvula gape nas águas, mesmo em condições de baixa visibilidade.
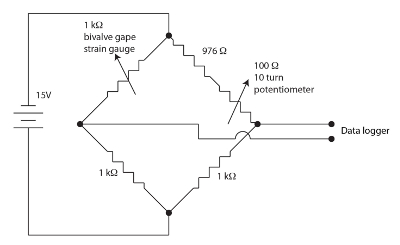
Figura 1. Ponte de Wheatstone para o aparelho de abertura da válvula. Clique aqui para ver uma versão maior desta figura.
Os calibre de tensão sensores para monitorar gape bivalves são filmes resistivos em um padrão de Meandro em um revestimento protetor de poliimida. Pequenas quantidades de estirpe modulam a resistência do sensor. Os bivalves flexiona o gage estirpe quando ele gapes causando uma mudança na resistência do sensor. Utilizamos uma ponte de Wheatstone anulação, equilibrada, para cada canal de bivalves, como mostrado na Figura 1 para medir a mudança na resistência do sensor. A ponte de Wheatstone é anulada pelo potenciômetro que permite um ganho bastante elevado ser empregado pelo datalogger. Uma ponte de Wheatstone é um método padrão para medir com precisão uma resistência desconhecida usando um rácio para uma resistência conhecida padrão e um voltímetro. A história desta técnica muito antiga é discutida em Ekelof (2001)60. 12 canais, cada um com sua própria ponte de Wheatstone e anulação potenciómetro, integrados a unidade Strain Gauge Monitor (SGM).
