Kemik bağ dokusu iki kısımdan oluşur bir türüdür: organik (hücreleri ve kollajen lifler) ve mineral (kalsiyum ve fosfat bileşikleri). Ana mineral kemikleri apatitler1bileşenleridir. Qafqaz yetkili hücreler kemik (dokusunu), diş (odontoblasts) ve kıkırdak (kondrosit) farklı türde hücre dışı matriks (ECM) proteinler üreten ve matris bırakmadan Qafqaz ilk adımları düzenleyen veziküller (MVs) (Şekil 1). MVs kalsiyum ve fosfat apatit çekirdekleşme kolaylaştırmak biriken ve daha sonra bağlamak kollajen2,3‘ e 100-300 nm çapında veziküller vardır. Sonra hücre dışı ortama apatitler serbest bırakmak için MVs parçalanır. Apatitler kollajen lifleri ile temas halinde büyümeye ve kemik matris oluşturmak devam. Qafqaz Pben ve Ca2 + ekstraselüler ortamda sürekli tedarik tarafından sürekli. Kısa bir süre önce bazı veriler bizim model4,5desteği. Yumuşak dokular fizyolojik koşullar altında mineralize değil. Ancak, ektopik kalsifikasyon vasküler kalsifikasyon3gibi patolojik şartlar altında meydana gelebilir. Osteoblast fenotip elde damar hücreleri apatitler çekirdekleşme teşvik ve kan damarlarının duvar medial ve intimal katmanlarındaki Qafqaz başlatmak MVs üretebilir. Ektopik kalsifikasyon beri normal endochondral Qafqaz3Qafqaz kemik hücreleri moleküler mekanizmaları anlama, benzer ve kondrosit bazı ipuçları vardır yumuşak dokuların ektopik önkol kemiğinde kireçlenme. sağlamalıdır kurdu.
İskelet doku gelişimi çeşitli enzimler, büyüme faktörleri ve rehberleri veya Qafqaz inhibitörleri tarafından düzenlenir. Uzlaşmaz eylem doku ettiren alkalen fosfataz (TNAP) (Şekil 1) ve ectonucleotide pyrophosphatase/fosfodiesteraz ben (NPP1), ankyrin (ANK), ile birlikte inorganik pirofosfat (PPben) konsantrasyonu kontrol eder 6. PPben, HA oluşumu, güçlü bir inhibitörü TNAP; hidrolize NPP1 nükleotit trifosfatlar arasında ANK PPben için ECM hücreden verir iken PPben oluşturmak üzere hidrolize. Pi/ÜFE oranı apatit oluşumu7,8 olası patolojik sonuçları9ile düzenleyen.
MV membran çekirdekleşme sürecinde (Şekil 1) kalsiyum ve fosfat MVs içinde yağış ilk kolaylaştırmak iyon taşıma proteinlerde zenginleştirilmiştir. Fosfat ışınlama 1 (çukur) MVs10,11perivesicular uzayda oluşturulan Pben dahil etmek için yardımcı olur. Annexins bağlama ve Ca2 + ulaşım ve Qafqaz MV lümen12,13‘ te başlattığı biyofiziksel süreç içinde tutulabilir. Biz daha önce apatit ECM14,15dakika sonra onun yayılma önce MV içinde iç çekirdekleşme intrasitoplazmik keseleri içinde Qafqaz için önerilen hipotez, iyilik. Vitro modelleme indüksiyon Ca2 +/Pben kompleksleri oluşumu PS ve AnxA516yapılan proteoliposomes doğruladı. Bu o birikimi gösteriyor olabilir sahip Ca2 +, Pben, lipid sallar, microvilli gibi membranesrepresent (NC) çekirdekleşme özünü apatit MVs. Annexins ve TNAP içinde AnxA5 ve PS kompleksleri ayrıca kollajen bağlayıcı kapasiteleri MVs kollajen lifleri boyunca koyarak ve ECM Qafqaz yayılmasını teşvik edici olarak yararlı olabilir. Fetuin A ve osteopontin (OPN)17, Qafqaz kolajen iskele üzerinde yayılmasını yavaşlatmak apatit oluşumu inhibitörleri olarak bilinir. Çekirdekleşme ve yayılma farklı olaylar, ikincisi, önceki eski vardır ve her ikisi de patolojik Qafqaz işlemi için uygun olmayabilir.
Kalsiyum fosfat kompleksleri Kimya fizyolojik Qafqaz ve ektopik kalsifikasyon nasıl değişebilir keşfetmek için hücreleri tarafından üretilen mineraller tanımlamak gereklidir. Apatitler kalsiyum ve fosfat genel kristal birim hücre formülü Ca10(PO4)6X2ile mineraller içeren bir grup, burada X = Cl, F, OH. 18aşağıdaki gibi sınıflandırılır: fluorapatite (FA) Ca10(PO4)6F2, chlorapatite (CA) Ca10(PO4)6Cl2 ve hidroksiapatit (HA) Ca10(PO4 )6(OH)2.
Her hücre kültürünü Qafqaz ayrı bir profil sergileyen bu yana osteoblast hücre hatları seçimi mineral oluşumu ikna etmek için çok önemlidir. Bu raporda, Qafqaz iki insan hücre modelleri tarafından mineraller çekirdekleşme karşılaştırıldığında: osteoblastik hFOB 1.19 hücreleri ve SAO’lar-2 osteosarkom hücreleri. Osteosarkom kaynaklı hücreler osteoblastik modelleri olarak yaygın olarak kullanılır ve farklılaşmamış insan fetal hFOB hücreler yaygın olarak normal osteoblastik bir model olarak kullanılır iken SAO’lar-2 hücreleri en olgun osteoblastik karakter19 korunmuş farklılaşma20. Qafqaz profillerini farklı yöntemlerle analiz edildi: Alizarin Red-S (AR-S) boyama, ultraviyole (UV) ışık görselleştirme, transmisyon elektron mikroskobu (TEM) görüntü, enerji dağıtıcı x-ışını microanalysis (EDX) Nefelometri ve iyon eşleme. TEM-EDX avantaj önceki çalışmalarda kullanılan alternatif teknikleri bu apatit kristallerinin4,5,21‘ iyon değiştirme nicel ve nitel sonuçlar veren şeydir. TEM-EDX kullanarak genel amacı görüntüleme ve miktar hücreler farklı türdeki çeşitli mineraller Ca, F ve Cl iyonu dağıtım Qafqaz sürecin farklı aşamalarında için basit bir yöntem bulmak oldu. Bu yöntem başarılı bir şekilde, örneğin, coexisting kimyasallar ile çinko nano tanecikleri ve Sucul organizmalar22kombine etkileri etkileşimi izlemek için kullanılmıştır. Başka bir çalışmada, bir bakır photocatalyst sulu çözüm titanyum malzemeleri hakkında kapsamlı İndüktif Eşleşmiş Plazma optik emisyon spektrometresi (ICP-OES), N2 physisorption (bahis), ile karakterize edildi XRD, UV-vis DRS, FT-IR, Raman Spektroskopi, TEM-EDX ve fotoelektrokimyasal ölçümler23. Amacımız kökeni ve veziküller ve mineraller Qafqaz kemik farklılaşma sırasında kontrol mekanizmasını anlamak için iki hücre satırı özelliklerini karşılaştırmak için oldu.
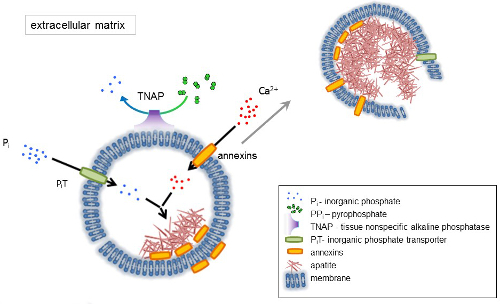
Resim 1 . Qafqaz kemik hücreleri içeren hücre dışı matriks (ECM) protein sentezi ve membran matris veziküller (MVs) sürümünden ilk adımların düzenini. MVs kalsiyum kalsiyum bağlayıcı proteinler, annexins ve inorganik fosfat ışınlama (çukur) eylem yoluyla hangi dephosphorylates doku non-spesifik alkalen fosfataz (TNAP), etkinlik tarafından takip fosfat eylem yoluyla birikir PPben böylece apatit çekirdekleşme kolaylaştırmak Pben. O zaman, MVs parçalanır ve hücre dışı ortama apatitler bırakın. Qafqaz Pben ve Ca2 + hücre dışı orta4,5sabit tedarik tarafından sürekli. Bu rakam daha büyük bir versiyonunu görüntülemek için buraya tıklayınız.
