Using the reprogramming strategy outlined above and in Figure 1B, we generated iCMs with approximately 70% of cells expressing cardiac Troponin T and approximately 55% of cells expressing cardiac α-actinin, quantified by flow cytometry at Day 9 following transduction of GHMT2m (Figure 2A and B). Additionally, the majority of cells express cardiac Troponin T, Troponin I, and cardiac α-actinin as well as the gap junction marker Connexin 43 at Day 14 following transduction (Figure 2C and D). Imaging with higher magnification reveals well-defined sarcomere structure formation and gap junctions forming between neighboring cells (white arrowheads, Figure 2D). Furthermore, spontaneous contraction and calcium transients indicate functionality of iCMs (Figure 3A-C and Movie 1).
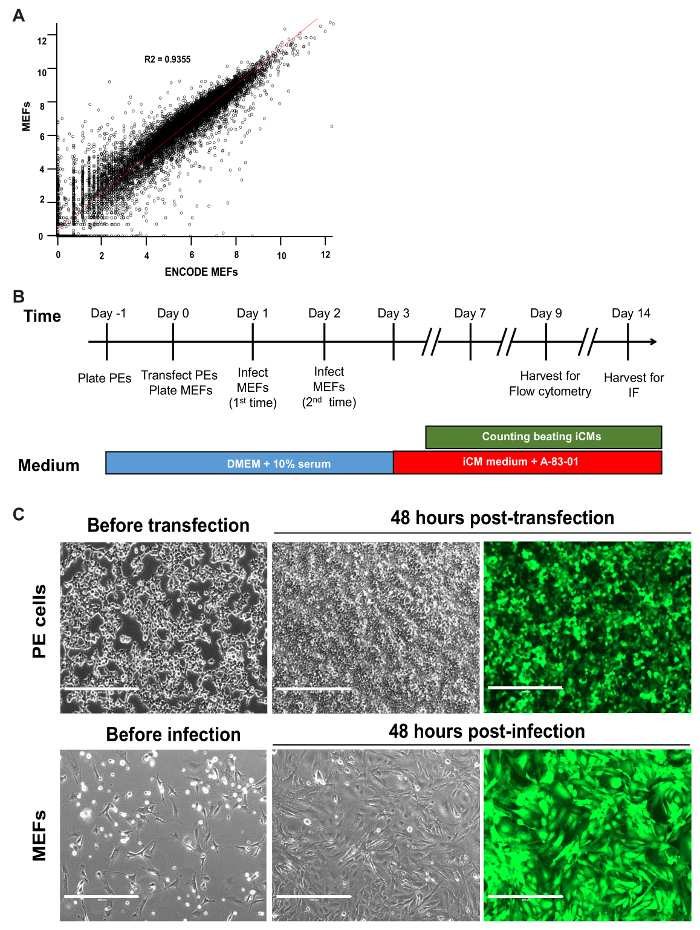
Figure 1: Timeline of Reprogramming and Optimal Plating of Cells. (A) Scatterplot of log-transformed RNA-seq data for MEFs generated in our lab versus ENCODE MEFs. The R2 value and linear regression line are shown. (B) Schematic of all critical steps in the GHMT2m-mediated reprogramming of MEFs. (C) PE cells at 70-80% confluence prior to transfection (top row, left panel) and GFP signal 48 hours post-transfection to indicate transfection efficiency (top row, middle and right panels). MEFs seeded sparsely prior to infection to prevent overcrowding over the time period of reprogramming (bottom row, left panel) and GFP signal 48 hours post-infection to indicate infection efficiency (bottom row, middle and right panels). Scale bar = 400 µM. Please click here to view a larger version of this figure.
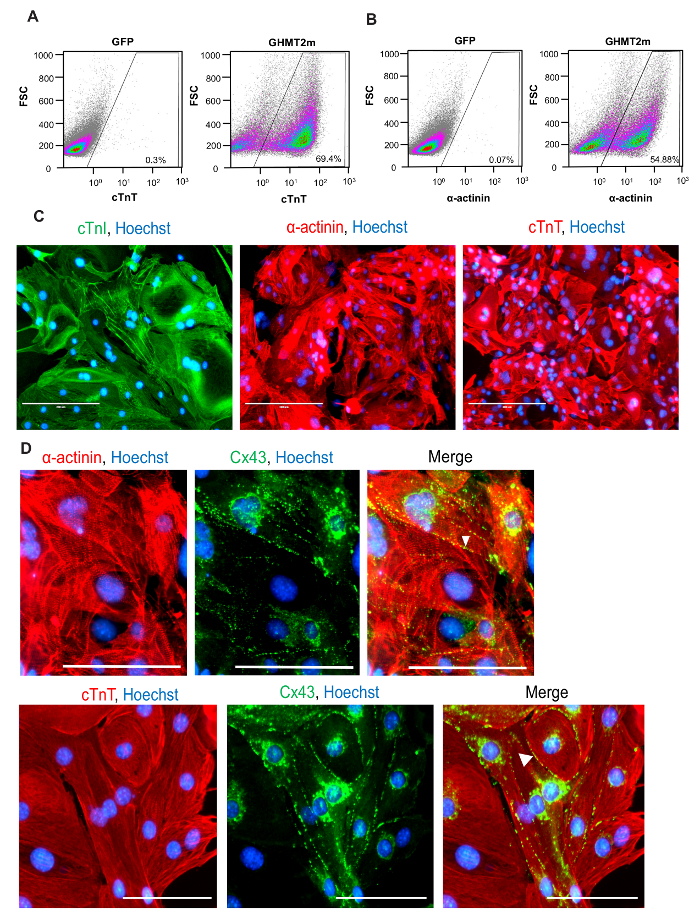
Figure 2: Quantification and Characterization of Cardiac Protein Expression in iCMs. (A and B). Representative flow cytometry for cardiac Troponin T (A) and α-actinin (B) at Day 9 following GHMT2m transduction, n = 3. (C) ICC staining for cardiac markers at Day 14 following GHMT2m transduction. Green: cardiac Troponin I, Red: cardiac Troponin T (middle panel) and α-actinin (right panel), and Blue: Hoechst staining for nuclei. Representative images (n = 3). Scale bar = 200 µM (D) High magnification imagines of sarcomere structure (Red: cardiac α-actinin (top row) and cardiac Troponin T (bottom row)) and expression of gap junction protein Connexin 43 (green). White arrowheads indicate gap junctions formed between neighboring cells. Representative images (n = 3). Scale bar = 100 µM. Please click here to view a larger version of this figure.
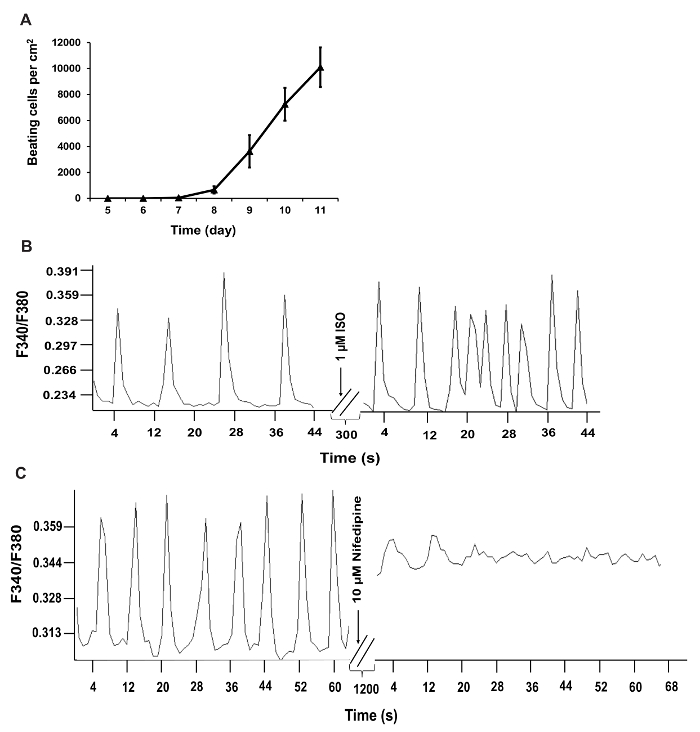
Figure 3: Functional Quantification of iCMs. (A) Time course of beating cell counts following GHMT2m transduction. Beating cells were counted by eye and cell counts from 10 fields per dish over 3 dishes per experiment were averaged and included in panel A. (B and C) Recorded calcium transients of iCMs. Calcium transient frequency is altered in iCMs following treatment with 1 µM Isoproterenol or 10 µM nifedipine. F340/F380, the ratio of fluorescence intensity at 340 and 380 nm. Please click here to view a larger version of this figure.
Movie 1: Beating iCMs. Movie showing beating iCMs at Day 12 in vitro. Please click here to view this video. (Right-click to download.)
| iCM Media | ||
| Name of Reagent | Volume (mL) | Final Concentration |
| DMEM High Glucose | 320 | |
| Medium 199 | 80 | |
| Fetal Bovine Serum | 50 | 10% |
| Donor Horse Serum | 25 | 5% |
| MEM Essential Amino Acids, 50x | 10 | 1x |
| Sodium Pyruvate Solution, 100x | 5 | 1x |
| MEM Non-Essential Amino Acids, 100x | 5 | 1x |
| MEM Vitamin Solution, 100x | 5 | 1x |
| Insulin-Transferrin-Selenium, 100x | 5 | 1x |
| B27, 50x | 10 | 1x |
| Penicilin-Streptomycin | 5.5 | 1.1% |
| L-Glutamine supplement | 5.5 | 1.1% |
| PE Media | ||
| Name of Reagent | Volume (mL) | Final Concentration |
| DMEM High Glucose | 450 | |
| Fetal Bovine Serum | 50 | 10% |
| Penicilin-Streptomycin | 5.5 | 1.1% |
| L-Glutamine supplement | 5.5 | 1.1% |
| Blasticidin-HCl – 10mg/mL | 0.5 | 10 µg/mL |
| Puromycin dihydrochloride | 0.05 | 1 µg/mL |
| Growth Media | ||
| Name of Reagent/ Equipment | Volume (mL) | Final Concentration |
| DMEM High Glucose | 450 | |
| Fetal Bovine Serum | 50 | 10% |
| Penicilin-Streptomycin | 5.5 | 1.1% |
| L-Glutamine supplement | 5.5 | 1.1% |
Table 1: Culture Media. Recipes for culture medium used in this protocol.
| PE Cells | ||||||
| Cell culture dish | Surface Area (cm2) | Seeding Density (cells) | Media Volume (mL) | Total DNA per transfection (µg) | Transfection reagent (µL) | Opti-MEM (µL) |
| 15 cm | 176.7 | 11.25 x 106 | 20 | 36 | 108 | 1080 |
| 10 cm | 78.5 | 5 x 106 | 10 | 12 | 36 | 360 |
| 60 mm | 28.2 | 1.7 x 106 | 4 | 4 | 12 | 120 |
| 6 well plate | 9 | 0.54 x 106 | 2 | 1.3 | 3.9 | 39 |
| 12 well plate | 4 | 0.24 x 106 | 1 | 0.6 | 1.8 | 18 |
| 24 well plate | 2 | 0.12 x 106 | 0.5 | 0.3 | 0.9 | 9 |
| MEFs | ||||||
| Cell culture dish | Surface Area (cm2) | Seeding Density (cells) | Media Volume (mL) | Hexadimethrine bromide (µL) | ||
| 15 cm | 176.7 | 1.35 x 106 | 20 | 12 | ||
| 10 cm | 78.5 | 0.6 x 106 | 10 | 6 | ||
| 60 mm | 28.2 | 0.2 x 106 | 4 | 2.4 | ||
| 6 well plate | 9 | 0.1 x 106 | 2 | 1.2 | ||
| 12 well plate | 4 | 0.4 x 105 | 1 | 0.6 | ||
| 24 well plate | 2 | 0.2 x 105 | 0.5 | 0.3 | ||
Table 2: Seeding Densities for Reprogramming. Table of seeding densities for PE cells and MEFs for common cell culture dish formats.
