Фотосинтетических организмов, таких как диатомовые водоросли должны справиться с постоянно меняющейся освещенности и реагировать с адаптационного сложные механизмы, которые поддерживать высокую эффективность фотосинтеза и защищать от фото окислительных повреждений, вызванных чрезмерным свет. Основной свет защитные процесс в фотосинтетические eukaryotes является высокой энергии закалочные (qE) поглощается света, который возникает как главный вклад не фотохимического тушения (ЕНОЛ) под легкий стресс условия1,2 ,3. Света уборки антенные комплексы (LHC) участвуют в регуляции пути передачи энергии возбуждения. В ответ на высокий свет индуцированной низкий рН в просвете хлоропластов, Антенные переключатели системы от света уборки тушения состояние. Этой энергии диссипативных государство защищает фотосистемы (PS) и других комплексов в тилакоидной мембране от фото окисления. В фотосинтетической эукариот qE индуцируется обычно двух факторов1,2,3. Одним из факторов является специализированным свет уборки белок, который реагирует с низким рН. ОВО белка индуцирует дЕ в высших растений4. LhcSRs5, модулированные деятельности ОВО, побудить дЕ в зеленых водорослей6. Диатомовые водоросли обладают Lhcx подобных белков, структурно связанные с LHCSRs7,8,9,10.
Второй фактор qE является цикл Ксантофилл где преобразован в форму фото защитные деэпоксидации и вернулась эпоксидирования каротиноиды антенны. В растениях и зеленые водоросли виолаксантина преобразуется в зеаксантин. В диатомовых водорослей diadinoxanthin преобразуется в diatoxanthin, который затем коррелирует с степень ЕНОЛ11. Диатомовые водоросли света уборки антенна обладает некоторые особенности, хотя это эволюционный, относящиеся к заводе и водорослей LHCs. Переход от света, заготовки для фото защиты чрезвычайно быстрый и ЕНОЛ способности выше по сравнению с растений12. Это может быть одной из причин, почему диатомеи очень успешно в различных экологических ниш в пути, что они отвечают за до 45% океанических чистой первичной продукции13. Таким образом диатомовые водоросли света уборки систем являются интересным объектом исследований фотосинтеза.
Диатомовые водоросли, как ориентированных видов Cyclotella meneghiniana, обладают тилакоидов внутренние свет заготовки систем, названный пигменты они связывают – фукоксантин, хлорофилла (ХЛ) a и c, поэтому FCP. света уборки белков, например FCPs, являются Встроенный в системе мембрана тилакоида состоит из нескольких слоев мембраны. Диатомовые водоросли образуют полосы три тилакоидов. Этот комплекс положение делает его трудным для их изучения на молекулярном уровне в тилакоидной мембраны. Кроме того многие компоненты способствуют регуляции света уборки (см. выше). Таким образом во многих подходов, комплексы были изолированы от мембраны, используя мягкий моющих средств, таких как н додецил β-D-maltopyranoside (β-DDM), который солюбилизировать последнего мембраны, но сохраняют FCP комплексов. Многие спектроскопические исследования были проведены с использованием растворимых FCP расследовать внутримолекулярной энергии передачи14,,1516,17. Однако этот бывший подход был ограничен, поскольку регулирование передачи энергии экситонных взаимодействие с другими антенные комплексы или фотосистемы. Следовательно, эти виды исследований не могут осуществляться с растворимых комплексов потому, что взаимодействие между комплексами теряется.
Важной особенностью в положении антенна является «молекулярная вытеснения» антенны и фотосистем в тилакоидной мембраны18. Ранее, простой подход было проведено, чтобы имитировать этот эффект в пробирке. Был удален моющих средств, что приводит к случайным агрегации антенных комплексов. Хотя некоторые разумные данных было получено17,этот подход19, моющего средства удаления не отражает ситуацию в естественных условиях и имеет некоторые ограничения, поскольку комплексы не взаимодействующих в их регулярных третичный структура.
Использование липосомы преодолевает несколько бывших ограничений. Третичная структура по-прежнему полностью нетронутыми. Липосомы мембраны обеспечивает квази родной среде для антенных комплексов. Мембрана отделяет внутри липосом из внешней среды. Этими средствами липосомы предоставляют две реакции отсеков для исследований Ион и pH градиентов, а также транспортных процессов. Кроме того параметры экспериментальной системы могут контролироваться более легко исследований в тилакоидной мембраны. Уже показано, что липосомы были отличным инструментом для изучения фотосинтетической комплексов. Основное внимание в прошлом был на заводе LHC, где влияние изменения липидного состава был протестирован на LHC II20. В других подходов протеин протеина взаимодействия между различными LHC II были исследованы21. Кроме того, некоторые в зеленых водорослей были проведены исследования описывают, спонтанное кластеризации между LHC22. Учитывая важность диатомовых водорослей для водных экосистем относительно немногие исследования проводились с антенные комплексы диатомовых водорослей. Два исследования исследованы антенные комплексы ориентированных Cyclotella meneghiniana, где были показаны кластеризации FCP антенна23 и оперативности FCP электрохимических градиентов24 . Таким образом липосомы являются отличным инструментом для изучения взаимодействия диатомовых антенн и их регулирования в условиях почти родной. Липосомы универсальны, поскольку многие условия, такие как липидного состава, липосомы размер, плотность белка и окружающей водной фазе может контролироваться. Кроме того этот метод требует низкого количества образцов. Экспериментальная система является надежной и высокую воспроизводимость. Изолированность липосомы позволяет для изучения рН и Ион градиенты, которые являются важными факторами в регулировании антенных комплексов.
Здесь мы описываем изоляции FCP антенных комплексов с C. meneghiniana и их включения в липосомах с естественным тилакоидов липидный состав. Кроме того мы предоставляем образцовые данные для спектральные характеристики растворимых FCP и сравнить их с FCP в липосомах. Метод обобщает знания и стандартизованных протоколов, полученные от улучшения Гундерманн и бушель 201223, Натали et al. 201622и24Ахмад и Дицеля 2017.
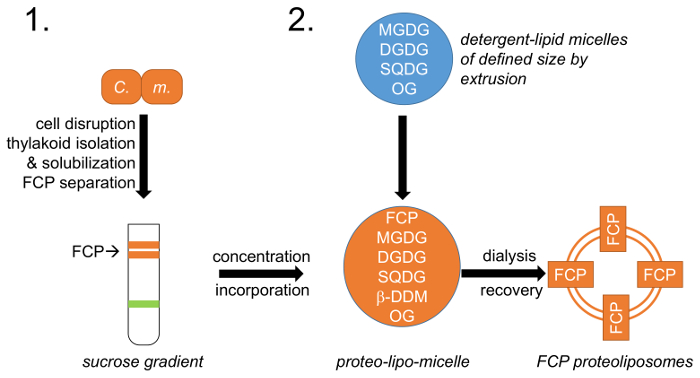
Рисунок 1: Схематическое представление рабочего процесса. (1) относится к пункту 1, который описывает рост клеток, нарушения и изоляции тилакоид с following FCP разделения на градиенты плотности сахарозы; C. м. –Cyclotella meneghiniana клетки. (2) приготовления смеси натуральных тилакоидов липидов (MGDG, DGDG и SQDG) описано в пункте 2 и создание мицелл липидов-моющего средства с octylglycoside (ОГ). Размер определенный липид мицеллы достигается методом экструзии, с использованием определенной поры диаметром мембраны. FCP и липидов мицеллы унифицированы на предопределенные липидов: соотношение белка и моющих средств, ОГ и β-DDM будут удалены через контролируемые диализа, образуя FCP proteoliposomes. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
