Fotosyntetiska organismer såsom kiselalger måste klara av ständigt föränderliga ljusförhållanden och svara med sofistikerade acklimatisering mekanismer som upprätthålla hög fotosyntetiska effektivitet och skydda från foto-oxidativ skada som orsakas av överdriven ljuset. En större tända-skyddande process i photosynthetic eukaryota organismer är hög energi släcka (qE) absorberas ljus som uppstår som den främsta bidragen till den icke-fotokemiska släcka (NPQ) under lätta stress villkor1,2 ,3. De lätta skörda antenn komplex (LHC) är involverade i regleringen av magnetiseringen energi överföring vägar. Svar på höga ljus inducerad lågt pH i kloroplast lumen, växlarna antenn system från ljuset skörd staten att släcka staten. Denna energi dissipativa staten skyddar fotosystem (PS) och andra komplex i tylakoida membran från photo-oxidation. I photosynthetic eukaryota organismer framkallas qE oftast av två faktorer1,2,3. En faktor är specialiserade ljuset skörd protein som svarar på den lågt pH. Proteinet PsbS inducerar qE i högre växter4. LhcSRs5, moduleras av PsbS aktivitet, framkalla qE i grönalger6. Kiselalger äger Lhcx-liknande proteiner som strukturellt besläktade LHCSRs7,8,9,10.
Den andra faktorn qE är xanthofyll cykeln där karotenoider av antennen konverteras till en foto-skyddande form av de-epoxidering och återgått av epoxidering. I växter och gröna alger konverteras violaxanthin till zeaxantin. I kiselalger omvandlas diadinoxanthin till diatoxanthin, som sedan korrelerar med omfattningen av NPQ11. Kiselalger ljuset skörd antenn besitter vissa egenheter även om det är evolutionärt relaterade till växter och alger LHCs. Övergången från ljus skörd till foto-skydd är oerhört snabb och NPQ kapacitet är högre jämfört med växter12. Detta kan vara en orsak varför kiselalger är mycket framgångsrika i olika ekologiska nischer på ett sätt som de är ansvariga för upp till 45% av den oceaniska netto primärproduktion13. Kiselalger ljus skörd system är därför ett intressant objekt av photosynthesis forskning.
Kiselalger, liknar centrerad arten Cyclotella meneghiniana, äger tylakoida inneboende ljus skörd system uppkallad efter pigment de binda – fucoxanthin, klorofyll (chl) a och c, därav FCP. ljus skörd proteiner, såsom FCPs, är inbäddad i tylakoida membran systemet bestående av flera membran lager. Kiselalger bilda band av tre thylakoids. Detta komplex situationen gör det svårt att studera dem på molekylär nivå i tylakoida membran. Dessutom bidrar många komponenter till regleringen av ljus skörd (se ovan). Därför i många metoder isolerades komplexen från membranet använder mild rengöringsmedel, till exempel n-Dodecyl-β-D-maltopyranoside (β-DDM), som solubilize membranet men hålla FCP komplexen intakt. Många spektroskopiska studier utfördes med solubilized FCP för att undersöka intramolekylära energi överföring14,15,16,17. Detta före detta tillvägagångssätt var dock begränsad eftersom regleringen av energiöverföringen behöver excitonic interaktion med andra antenn komplex eller fotosystem. Därför, dessa typer av studier inte kan utföras med solubilized komplex eftersom interaktionen mellan komplex går förlorad.
Ett viktigt inslag i antenn förordning är ”molekylär trängsel” antenn och fotosystem i tylakoida membran18. Tidigare, en enkel strategi genomfördes för att simulera effekten in vitro-. Tvättmedlet togs bort, vilket leder till slumpmässiga aggregering av antenn komplex. Även om vissa rimliga uppgifter erhölls genom detta tillvägagångssätt17,19, tvättmedel avlägsnande återspeglar inte den situation i vivo och har vissa begränsningar eftersom komplexen inte interagerar i sin regelbundna tertiär struktur.
Användning av liposomer övervinner flera av de tidigare begränsningarna. Tertiär struktur är fortfarande helt intakt. Liposom membranet ger en kvasi infödda miljö för antenn komplexen. Membranet separerar insidan av Liposom från den yttre miljön. Av dessa medel ger liposomer två reaktion fack för studier av ion och pH lutningar samt när det gäller processer. Ytterligare, parametrarna för experimentell systemet kan kontrolleras lättare för studier i tylakoida membran. Liposomer har redan visat sig vara ett utmärkt verktyg att studera fotosyntetiska komplex. Ett stort fokus i förflutnan var på anläggning LHC där effekten av förändrade lipid sammansättning testades på LHC II20. I andra metoder, protein-protein interaktioner mellan olika LHC II var undersökta21. Också, vissa studier i grönalger genomfördes som beskriver spontana klustring mellan LHC22. Som beaktar betydelsen av kiselalger för akvatiska ekosystem, relativt få studier utfördes med antenn komplex av kiselalger. Två studier undersökt antenn komplexen av den centrerad Cyclotella meneghiniana, var den klustring av FCP antenn23 och lyhördhet av FCP till elektrokemiska övertoningar24 visades. Liposomer är alltså ett utmärkt verktyg att studera kiselalger antenner och deras interaktion och förordning i nästan inhemska förhållanden. Liposomer är mångsidig eftersom många förhållanden såsom lipid sammansättning, Liposom storlek, protein densitet och kringliggande vattenfasen kan styras. Metoden kräver dessutom låga mängder prover. Det experimentella systemet är robusta och mycket reproducerbara. Uppdelning i liposomer möjliggör studera pH och ion övertoningar, vilka är viktiga faktorer i regleringen av antenn komplex.
Här beskriver vi isolera FCP antenn komplex från C. meneghiniana och deras införlivande i liposomer med naturliga tylakoida lipid sammansättning. Också, vi tillhandahåller exemplariskt data för spektroskopisk karakterisering av solubilized FCP och jämföra dem med FCP i liposomer. Metoden sammanfattar kunskap och standardiserade protokoll erhålls från förbättringar av Gundermann och Büchel 201223, Natali et al. 201622och Ahmad och Dietzel 201724.
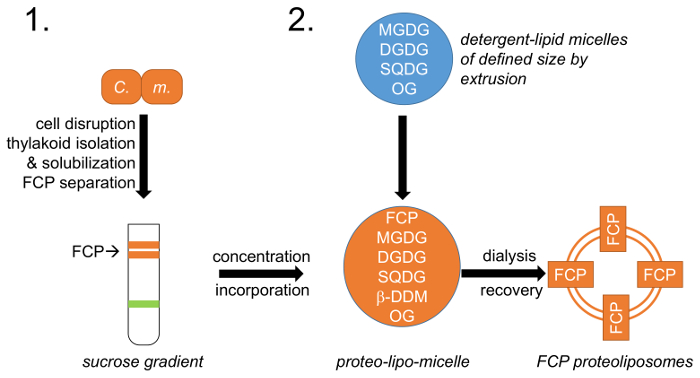
Figur 1: Schematisk bild av arbetsflödet. (1) refererar till punkt 1 och som beskriver celltillväxt, störningar och tylakoida isolering med följande FCP separation på sackaros täthetlutningar; C. m. –Cyclotella meneghiniana celler. (2) beredning av naturliga tylakoida lipid blandning (MGDG, DGDG och SQDG) beskrivs i punkt 2 och skapandet av lipid-tvättmedel miceller med octylglycoside (OG). En definierad lipid-micelle storlek uppnås genom extrudering med membran av definierade pordiameter. FCP och lipid-miceller är enhetlig på en fördefinierad lipid: förhållandet mellan protein och tvätt OG och β-DDM är borttagna via kontrollerade dialys bildar FCP proteoliposomes. Klicka här för att se en större version av denna siffra.
