Mouse model to isolate SMA+ MECs and pericytes
The established protocol allows for the isolation of two pure populations: MECs and pericytes from LGs and SMGs (see Table 1). These two types of cells have a different size and appearance. Microvascular pericytes, develop around the walls of capillaries (Figure 5A) and have a squared shape (Figure 5B), while MECs surround the LG secretory acini, have long processes and occupy a relatively large area (Figure 5A,B). The described procedure is based on genetic cell labeling of SMA+ in the TM-inducible SMACreErt2/+:Rosa26-TdTomatofl/fl mouse strain. Additonally, EpCAM antibodies allow the researchers to distinguish epithelial SMA+:EpCAM+ cells of ectodermal origin (MECs) and SMA+EpCAM– cells of endodermal origin (pericytes).
Preparation of single-cell suspension
The LG contains a filamentous extracellular matrix that must be digested thoroughly. The provided protocol allows preparation of a single cell solution for FACS analysis and further applications. The example of dissociated cells is shown in Figure 3.
MEC and pericyte isolation by FACS
To distinguish MECs from pericytes, single cells were stained with antibody to EpCAM, which detects only epithelial cells. The main population of cells was determined by forward and side scatter area gating (Figure 4A). Doublets were excluded by plotting the forward scatter area versus width and with the side scatter area versus width (Figure 4B). Dead cell exclusion was done via Ghost Red 780 (Viability Dye) (Figure 4C). An unlabeled control (Figure 4E), background control and antibody control labeled with a single primary antibody were used to determine the background noise (nonspecific antibody binding) and to establish proper compensation for optimum separation between signals (Figure 4D). Data analyses were performed by using FlowJo software.
MECs and pericytes were gated by DsRed labeling. DsRed+ dim (not shown) and DsRed+ bright cells within both MEC and pericyte cell populations were detected (Figure 4D). The brightness of labeled cells may depend on level of SMA expression or degree of reporter activation upon TM injection19. Only DS Red+ bright cell population were collected since only fully differentiated cells were required. The DS Red+ dim cell populations require further investigation.
Downstream applications
It is well known that MECs play an important contractile function in exocrine glands. Moreover, they are very plastic cells and have features of stem cells. Therefore, isolated MECs could be used in multiple applications. For example, cells can be cultured, used for RNA isolation or transplantation (Figure 1F)12,20,21,22.
| Parameter | Lacrimal gland | Submandibular gland |
| Number of mice per sample | 2 | 1 |
| Number of glands per sample | 4 | 2 |
| Dissection glands | Separate from parotid gland | Separate from sublingual gland |
| Concentration of collagenase per sample | 6 mg/2 mL | 9 mg/2 mL |
| Approximate cell number after enzymatic dissociation | 4×105-6×105 | 9×105-1.5×106 |
| Recovery step (see the section “Adult Mouse Lacrimal Gland Single-cell Dissociation”, point 15) | Re-suspend cells in 6 ml of recovery media | Re-suspend cells in 12 mL of recovery media |
| Volume of FACS buffer during antibody staining | 400 µL | 2 tubes by 400 µL; it is better to split the cells into two or three tubes that each tube has not more than 6×105 |
Table 1: Modifications of the protocol for isolation of cells from the submandibular gland (SMG). The table describes major modifications required to isolate MECs and pericytes from murine SMG in comparison with the procedure for murine LG.
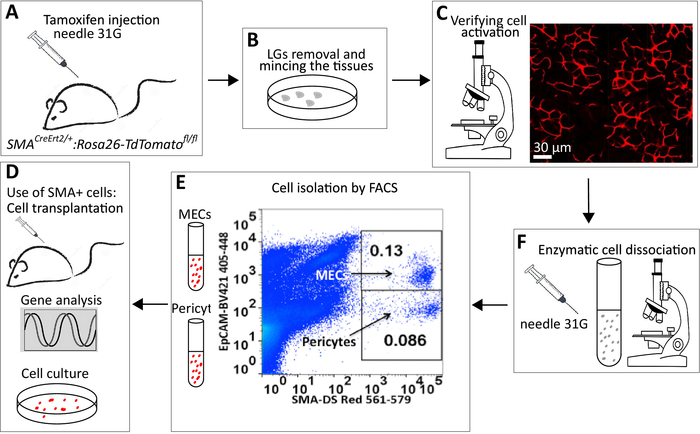
Figure 1: A schematic representation of the experiment. (A) IP injections of the SMACreErt2/+:Rosa26-TdTomatofl/flmice with TM. (B) Isolation and mincing of the LG or SMG. (C) Analysis of cell labeling using fluorescence microscope. (D) Multi-step enzymatic digestion to prepare a single-cell solution. It is critical to check digestion steps under a light microscope to ensure that cells are released from clusters. (E) Example of gating showing SMA+ bright DS Red+/EpCAM+ (MECs) and DS Red+bright/EpCAM- (pericytes). (F) Collected MECs and pericytes could be subjected to different downstream procedures including cell cultivation, RNA isolation and gene expression analysis and cell transplantation. Please click here to view a larger version of this figure.
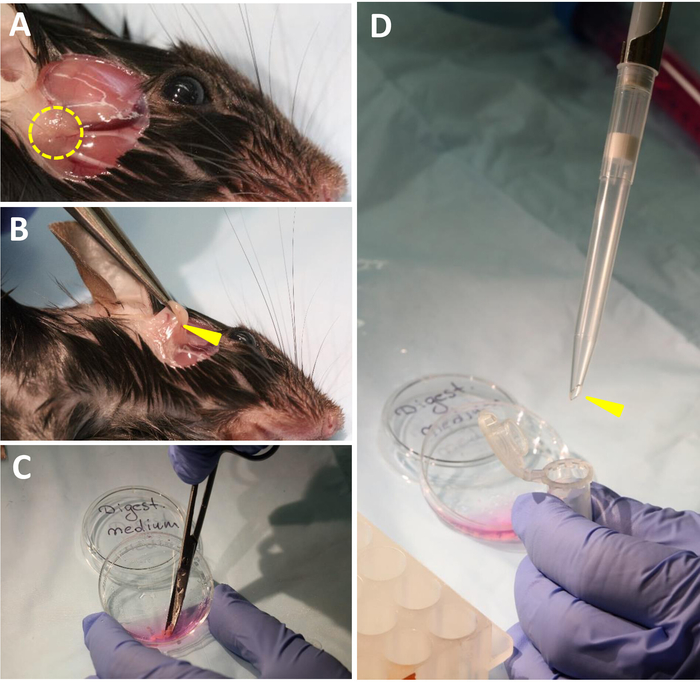
Figure 2: Critical steps of LG isolation/mincing. (A) Removal of skin between the eye and ear to dissect LG. Dashed yellow circle indicates LG location. (B) LG dissection. Yellow arrowhead points out area between lacrimal and parotid glands. (C) LG mincing in digestion medium using scissors with curved, blunt ends. (D) Transfer of minced tissue into 2 mL tube. Yellow arrowhead depicts wide bore size 1 ml tip required for transferring. Please click here to view a larger version of this figure.
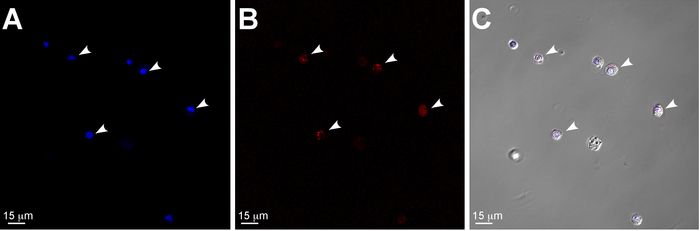
Figure 3: Confocal and differential interference contrast (DIC) images illustrating dissociated single cells from murine LG. (A-C) single cells isolated from two LGs of one 4 month old SMACreErt2/+:Rosa26-TdTomatofl/fl mouse. Nuclei are stained with DAPI (blue). White arrowheads denote SMA+ (DS Red) cells: MECs or pericytes. Scale bar = 15 μm. Please click here to view a larger version of this figure.
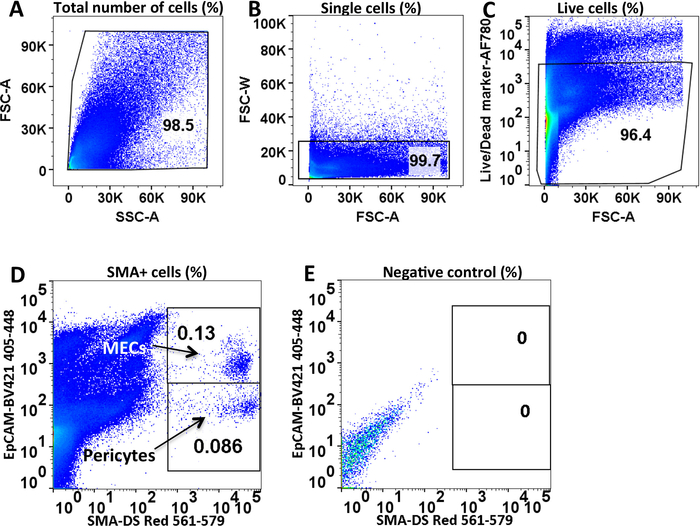
Figure 4: Identification of murine LG MECs and pericytes using FACS. (A) Determination of the main population of LG cells by forward and side scatter area gating. (B) Exclusion of doublets via forward scatter area versus width. (C) Dead cell exclusion via Ghost Red 780 (Viability Dye). (D) MEC (SMA+ bright/EpCAM+) and pericyte (SMA+ bright/EpCAM–) populations distinguished based on staining with EpCAM antibody. (E) Unlabeled control (cells from wild type mouse). On each plot the % of gated cells is provided. Please click here to view a larger version of this figure.
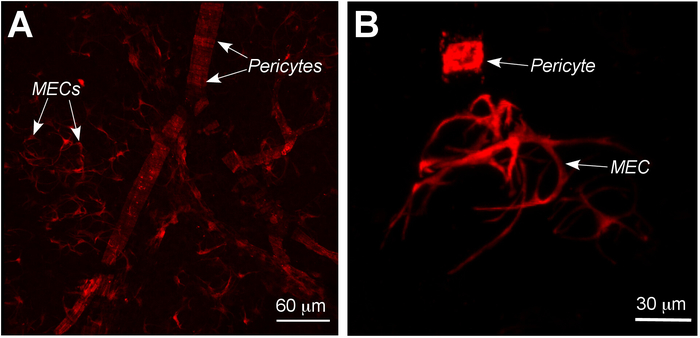
Figure 5: Confocal images showing difference in distribution and shape between MECs and pericytes isolated from murine LG. (A) Whole mount preparation of LG with labeled cells showing difference in distribution between MEC and pericytes. (B) The shape of MEC and pericyte is different. Pericyte is relatively small and has squired shape, whereas MEC is large and has irregular shape and several long processes. Arrowheads = MECs and pericytes. Please click here to view a larger version of this figure.
