Alcune specie reattive dell’ossigeno (ROS) sono in grado di ossidare doppi legami di carbonio di basi di DNA e alcuni carboni nella frazione desossribosio, generando basi ossidate e rottura del filamento del DNA1. Come una molecola carica negativamente ricca di atomi di azoto e ossigeno, il DNA è anche un bersaglio per i gruppi elettrofilici che covalentemente reagiscono con i siti nucleofili (azoto e ossigeno), dando prodotti che sono chiamati addotti del DNA2. Quindi, gli addotti del DNA e le basi del DNA ossidato sono esempi di lesioni del DNA che sono biomarcatori utili per la valutazione della tossicità di sostanze che sono elettrofiliche, generano elettrofili reattivi sulla biotrasformazione, o inducono lo stress ossidativo1, 2. il Anche se le basi di DNA modificate possono essere rimosse dal DNA per riparazione di escissione di base o nucleotide (BER o NER), l’induzione di uno squilibrio tra la generazione e la rimozione delle lesioni del DNA a favore della prima porta ad un aumento netto dei loro livelli nel lavoro straordinario del DNA3 . I risultati sono l’aumento dei tassi di mutazione del DNA, la riduzione dell’espressione genica e la diminuita attività proteica2,4,5,6,7, effetti che sono strettamente correlati alla sviluppo di malattie. Le mutazioni del DNA possono influenzare diverse funzioni cellulari, come la segnalazione cellulare, il ciclo cellulare, l’integrità del genoma, la stabilità dei telomeri, l’epigenoma, la struttura della cromatina, lo splicing dell’RNA, l’omeostasi proteica, il metabolismo, l’apoptosi e la differenziazione cellulare8 ,9. Le strategie per rallentare i tassi di mutazione cellulare e lo sviluppo di malattie croniche (ad esempio, cancro, malattie neurodegenerative) passano attraverso la conoscenza delle fonti di mutazione, tra cui le lesioni del DNA e le loro cause.
ROS ha generato in eccesso in modo endogeno, a causa di esposizione di inquinanti, infiammazione persistente, patofisiologia della malattia (ad esempio, diabete), ecc., sono cause importanti di danni biomolecola, tra cui il danno del DNA e del lipido1. Ad esempio, il radicale idrossile altamente reattivo (OH) formato da H2O2 riduzione degli ioni metallici di transizione (Fe2 +, cu+) ossida le basi del DNA, la porzione di zucchero del DNA e gli acidi grassi polinsaturi a diffusione controllata tassi di cambio10. Tra i 80 già caratterizzati nucleobasi ossidate3, l’uno più studiato è 8-oxo-7, 8-diidroguanina (8-oxoGua) o 8-oxo-7, 8-dihydro-2′-deossiguanosina (8-oxodGuo, Figura 1), una lesione che è in grado di indurre le trasversioni gt in cellule di mammifero10,11. È formata dall’ossidazione mono-elettronica della guanina, o da un attacco di ossigeno radicale idrossile o singollet di guanina nel DNA1. Gli acidi grassi polinsaturi sono altri importanti bersagli di ossidanti altamente reattivi, quali •Oh, che avviano il processo di perossidazione lipidica1,12. Dà origine a idroperossidi di acidi grassi che possono decomporre a aldeidi elettrofiliche ed epoxyaldeidi, come malondialdeide, 4-idrossi-2-nonenal, 2, 4-decadienal, 4,5-epossidico-(2E)-decenale, hexenal, Acrolein, crotonaldeide, che sono in grado di formare addotti mutageni del DNA esociclico, come malondialdeide-, propano-, o addotti di addotti1,12,13. L’addotti addotti 1,n2-addotti-2′-deoxyguanosine (1,n2-εdguo, Figura 1) e 1,n6-addotti-2′-deossiadenosina (1,n6-εdado, Figura 1 ) sono stati suggeriti come potenziali biomarcatori nella patofisiologia dell’infiammazione14,15.
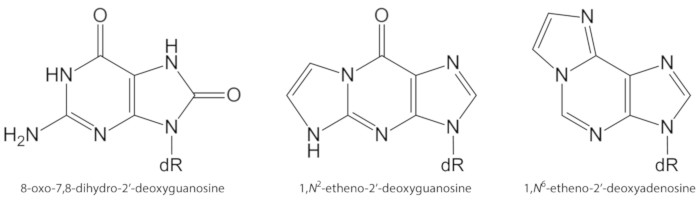
Figura 1. Strutture chimiche delle lesioni del DNA quantificate nel presente studio. dR = 2 ́-desossribosio. Questa cifra è stata modificata da Oliveira et al.34. Si prega di cliccare qui per visualizzare una versione più grande di questa cifra.
Gli studi condotti nei primi anni ottanta hanno permesso la rilevazione sensibile di 8-oxodGuo mediante cromatografia liquida ad alte prestazioni accoppiata al rilevamento elettrochimico (HPLC-ECD). Quantificazione di 8-oxodGuo da HPLC-ECD in diversi sistemi biologici sottoposti a condizioni ossidanti ha portato al riconoscimento di 8-oxodGuo come un biomarcatore di danno base ossidativamente indotto in DNA1,16. Sebbene robusto e che consenta la quantificazione di 8-oxodGuo nella gamma bassa fmol17, le misurazioni HPLC-ECD si basano sull’accuratezza del tempo di ritenzione dell’analita per l’identificazione dell’analita e sulla risoluzione della cromatografia per evitare interferenze di altri costituenti campione. Poiché la rilevazione elettrochimica richiede l’uso di sale (ad esempio fosfato di potassio, acetato di sodio) nella fase mobile, il mantenimento di condizioni analitiche adeguate necessita di routine di pulizia delle colonne e delle attrezzature.
In alternativa, l’uso dell’enzima di riparazione del DNA batterico formamidopirimidina DNA glicosilasi (FPG) e, successivamente, umana 8-oxoguanina glicosilasi 1 (hOGG1), per il rilevamento e la rimozione di 8-oxoGua dal DNA, è emerso come un modo per l’induzione del DNA alcalino labile Siti. I siti alcalini labili vengono convertiti in rotture del filamento del DNA e consentono la quantificazione indiretta molto elevata sensibile di 8-oxoGua mediante elettroforesi di gel monocellulare alcalina (“saggio Comet”). L’alta sensibilità e la realizzazione delle analisi senza la necessità di estrazione del DNA cellulare sono i principali vantaggi di questo tipo di saggio. Fornisce i livelli più bassi di stato stazionario di 8-oxoGua nel DNA, tipicamente 7-10 volte inferiori ai livelli ottenuti dai metodi bioanalitici basati su HPLC. Tuttavia, è una misura indiretta di 8-oxogua e alcuni inconvenienti sono la mancanza di specificità o l’efficienza sconosciuta degli enzimi di riparazione utilizzati1,16,18.
Gli immunodosaggi sono altri metodi utilizzati per la rilevazione di addotti di 8-oxoGua1 e di DNA esociclico, come 1,n6-dAdo e 1,n2-dGuo12. Nonostante la sensibilità, una carenza dell’uso di anticorpi per il rilevamento delle lesioni del DNA è la mancanza di specificità a causa della reattività incrociata ad altri componenti di campioni biologici, comprese le normali basi di DNA1,12. Gli addotti del DNA exociclico, compresi 1, n6-dado e 1, n2-dGuo, possono essere rilevati e quantificati da 32P-saggi di postetichettatura12. L’elevata sensibilità di 32P-postetichettatura consente l’uso di piccolissime quantità di DNA (ad es. 10 μg) per il rilevamento di circa 1 addotto per 1010 basi normali19. Tuttavia, l’uso di radio-chimiche, mancanza di specificità chimica e bassa precisione sono alcuni svantaggi19,20.
Una limitazione condivisa dei metodi citati sopra è la bassa selettività o specificità per la rilevazione delle molecole desiderate. In questo scenario, l’HPLC accoppiato alla spettrometria di massa tandem a ionizzazione elettrospray (HPLC-ESI-MS/MS e HPLC-MS3) si è evoluto come lo standard Gold per la quantificazione dei nucleosidi modificati nelle matrici biologiche, come il DNA, l’urina, il plasma e la saliva 1 il , 19 anni di , 20. i vantaggi dei metodi HPLC-ESI-MS/MS sono la sensibilità (tipicamente nella gamma bassa fmol) e l’elevata specificità fornita da i) la separazione cromatografica, II) il modello caratteristico e noto della frammentazione molecolare all’interno della massa Camera di collisione dello spettrometro, e III) la misurazione accurata del rapporto massa/carica selezionato (m/z) in modalità di monitoraggio a reazione multipla1,19. L’uso di standard interni con etichetta isotopica aggiunge il vantaggio delle correzioni per le perdite di molecola durante le fasi di arricchimento dell’idrolisi e dell’analita del DNA, nonché per le differenze di ionizzazione dell’analita tra i campioni. Aiuta anche a identificare il picco cromatografico corretto quando più di un picco è presente1,12,19,20.
Diversi metodi basati su HPLC-ESI-MS/MS sono stati utilizzati per la quantificazione di 8-oxodGuo, 1,N6-dAdo e 1,n2-dGuo nel DNA Estratto da diversi campioni biologici12,15,20 ,21,22,23,24,25,26,27,28,29 . Le particelle sottili (PM2,5) trasportano sostanze chimiche organiche e inorganiche, quali idrocarburi policiclici aromatici (IPA), Nitro-IPA, aldeidi, chetoni, acidi carbossilici, chinolini, metalli e ioni solubili in acqua, che possono indurre infiammazione e stress ossidativo, condizioni che favoriscono il verificarsi di biomolecola danni e malattia30,31,32,33. Qui presentiamo i metodi HPLC-ESI-MS/MS convalidati che sono stati applicati con successo per la quantificazione di 8-oxodGuo, 1,n6-dado e 1,n2-dGuo in polmone, fegato e DNA renale di un/J topi per la valutazione del effetti dell’esposizione ambientale PM 2,534.
