The protocol described in Figure 1A generated highly pure cardiomyocytes that acquire a ventricular/adult-like phenotype with time in culture. As assessed by immunofluorescence staining for the atrial and ventricular myosin regulatory light chain 2 isoforms (MLC2A and MLC2V, respectively), the majority of the cells generated by this protocol were MLC2A-positive at day 30 after induction of cardiac differentiation, while MLC2V was expressed in much lower amounts at the same time point (Figure 2A, top panels). As the time in culture increased (day 60 and 90), a complete switch of MLC2 isoforms (MLC2A to MLC2V) was observed (Figure 2A, bottom panels). In order to quantify the MLC2A and MLC2V-positive cells, flow cytometry analysis was performed. In accordance with the immunofluorescence results, the flow cytometry data demonstrated early stage (day 30) iPSC-CMs mostly expressing MLC2A (29.8% MLC2A + vs. 1.9% MLC2V +) (see Figure 2B, left panel), as compared to late stage (day 90) iPSC-CMs, which mostly expressed MLC2V (41.3% MLC2V + vs. 16.7% MLC2A +) (see Figure 2B, right panel). Because the expression pattern of MLC2A and MLC2V is known to be a hallmark of cardiac differentiation and maturation, these results suggest that prolonged culture time increases maturation of iPSC-CMs and that the majority of cells appear to be committed to the ventricular phenotype. We then assessed the time dependence of Ca2+ handling maturation. Ca2+ transient was measured in iPSC-CMs at the three differentiation times (day 30, 60, and 90), and compared to isolated adult rat cardiomyocytes (ARCMs). The Ca2+ amplitude was significantly increased in the iPSC-CMs at day 90 and was similar to the ARCMs (Figure 3A,B). The rate of Ca2+ reuptake (decay-tau) at day 90 was significantly faster compared to day 30 iPSC-CMs, and closer to ARCMs (Figure 3C). The effect of β-adrenergic stimulation on Ca2+ transients was further evaluated by treating the cells with 10 nM of isoproterenol (ISO) for 10 min at 37 °C. As observed in ARCMs, ISO significantly increased Ca2+ transient and accelerated the rate of Ca2+ reuptake in day 90 iPSC-CMs. No changes were observed in day 30 iPSC-CMs (Figure 3D-F). Interestingly, day 90 iPSC-CMs were able to follow increasing electrical stimulation (from 0.5−3 Hz), in a similar manner to that observed in ARCMs (Figure 4B). Taken together, these results show that iPSC-CMs derived from this method have similar characteristics to those seen in native CMs, specifically ventricular-like phenotypes, mature Ca2+ handling properties, and positive β-adrenergic responses.
Contaminating noncardiac cells may influence the functional maturation of human iPSC-CMs during differentiation17. Figure 4A shows a comparison between 3 month-old cells obtained from high efficiency differentiations (top panels, Video 1), robustly expressing the specific ventricular marker (MLC2V), and 3-month-old cells obtained from low efficiency differentiations (bottom panels, Video 2), showing iPSC-CMs mixed with alpha-smooth muscle actin (αSMA)-positive cells. Interestingly, functional analysis demonstrated altered Ca2+ transients in the mixed iPSC-CMs/non-CMs preparation compared to the iPSC-CMs from the high efficiency differentiation. In particular, the cells obtained from low efficiency differentiations exhibited very small Ca2+ amplitude and automaticity (Figure 4B, middle panel) compared to pure iPSC-CMs (Figure 4B, top panel) and ARVCs (Figure 4B, bottom panel). In addition, cells from low efficiency differentiations presented arrhythmic patterns (Figure 4C).
It is important to note that the iPSC-CMs shown in the top panel of Figure 4A also underwent metabolic selection, which is an important step to further enrich a population of iPSC-CMs that is already derived from a highly efficient differentiation. These data indicate that high efficiency cardiac differentiation protocols that generate high-quality CMs are necessary to accurately recapitulate a cardiac phenotype in vitro.
Figure 5 shows the Ca2+ amplitude variability throughout different areas of the CMs monolayer, plotted over a time course of 1,200 s. The Ca2+ amplitudes measured at the beginning of a stimulation frequency of 1 Hz (200 s) were highly variable and became more consistent as the stimulation continued (200−900 s). However, after a time period of 900 s the Ca2+ amplitude was considerably reduced. These data indicate that, when recording Ca2+ transients in iPSC-CMs, it is necessary to let the cells stabilize at 37 °C under constant stimulation for at least 200 s. Additionally, recordings have to be restricted to a specific time window (200−900 s) to ensure reproducible results.
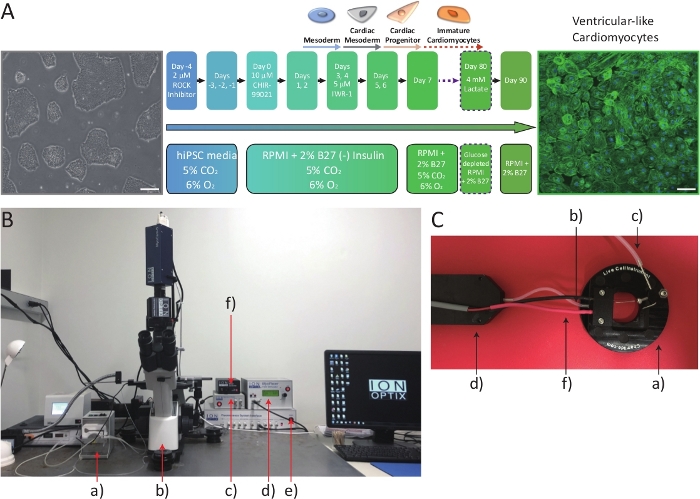
Figure 1: Overall schematic of iPCS-CMs preparation and Ca2+ transient instruments. (A) Schematic of cardiac differentiation protocol showing the developmental stages of differentiating iPSCs. Scale bar = 50 μm. (B) The calcium and contractility acquisition and analysis system. a) Peristalitic pump b) Inverted microscope c) Power source for the digital camera d) Electrical stimulator e) Fluorescence system interface f) Filter wheel controller (C) System chamber. a) System chamber body. b) Fluid inlet. c) Fluid outlet. d) Fluidic inline solution heater. f) Electrodes connecting the system chamber to the electrical stimulator. Please click here to view a larger version of this figure.
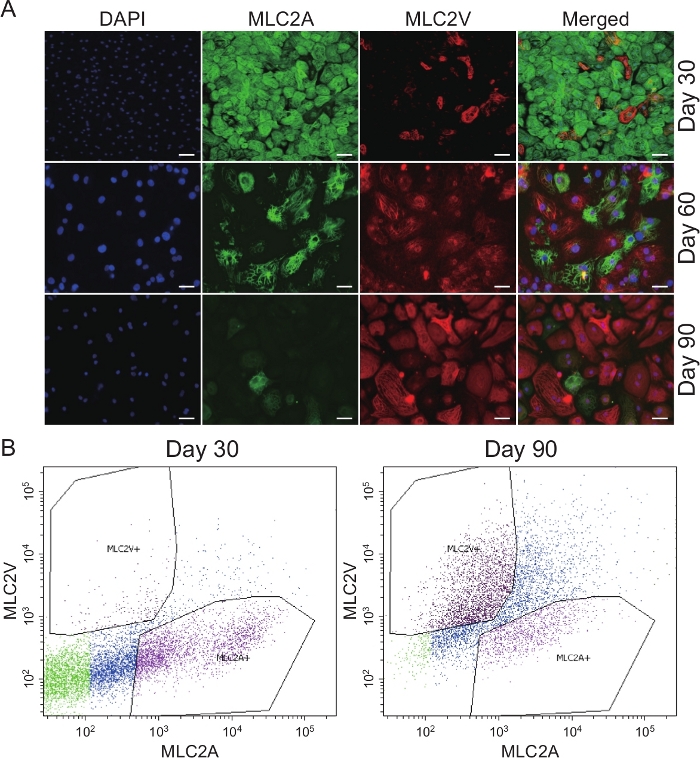
Figure 2: Comparison of MLC2A and MLC2V expression in iPSC-CMs at different days of differentiation. (A) Immunofluorescence staining of iPSC-CMs at day 30, 60, and 90 after differentiation induction for MLC2A and MLC2V. Scale bar = 20 μm. (B) Flow cytometry analysis of iPSC-CMs for MLC2V and MLC2A at 1 month and 3 months post differentiation. Please click here to view a larger version of this figure.
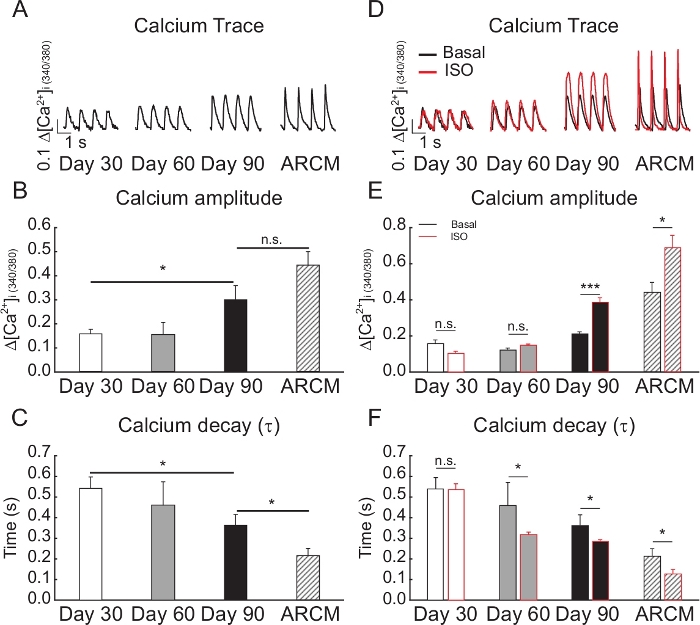
Figure 3: Comparison of iPSC-CMs properties at different days of differentiation. (A) Representative Ca2+ transient traces at different days of differentiation. (B–C) Average Ca2+ amplitude and Ca2+ decay elicited during stimulation at 1 Hz. (D–F) Effect of isoproterenol (ISO, 10 nM) treatment in iPSC-CMs at different days of differentiation. ARCMs indicate isolated rat adult cardiomyocytes. N = 70−100 areas; *p < 0.05; ***p < 0.001, as determined by Student's t-test. Data are represented as Mean ± S.E.M. Please click here to view a larger version of this figure.
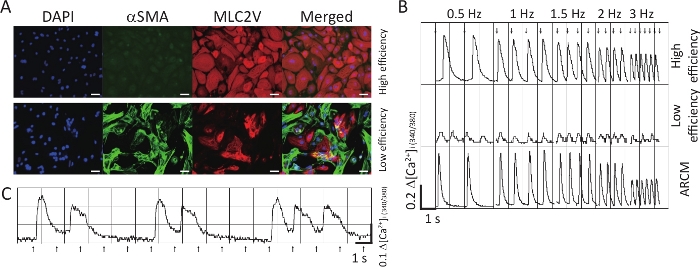
Figure 4: Comparison between high efficiency and low efficiency differentiations of iPSC-CMs. (A) Immunofluorescence staining of iPSC-CMs for αSMA and MLC2V. Scale bar = 20 μm. (B) Representative traces showing Ca2+ transients from high efficiency (above) and low efficiency (middle) differentiations, and isolated rat adult cardiomyocytes (below). The myocytes were stimulated at 0.5 Hz, 1 Hz, 1.5 Hz, 2 Hz, and 3 Hz. (C) Representative trace showing an arrhythmic pattern from a bad differentiation. Arrows indicate point pacing. ARCMs indicates isolated rat adult cardiomyocytes. Please click here to view a larger version of this figure.
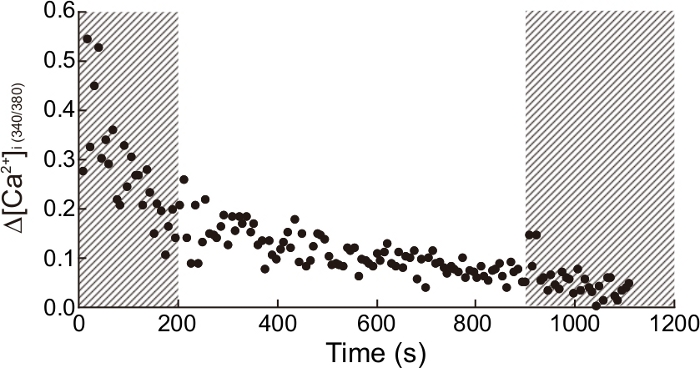
Figure 5: Time-dependent Ca2+ amplitude from a single coverslip. Ca2+ amplitudes from different areas in a coverslip were plotted in a time-dependent manner. Non-recommended time periods to measure Ca2+ transient are indicated in the dashed areas (<200 s or >900 s). Please click here to view a larger version of this figure.

Video 1: Representative video showing an example of a high efficiency differentiation. Please click here to download this video.
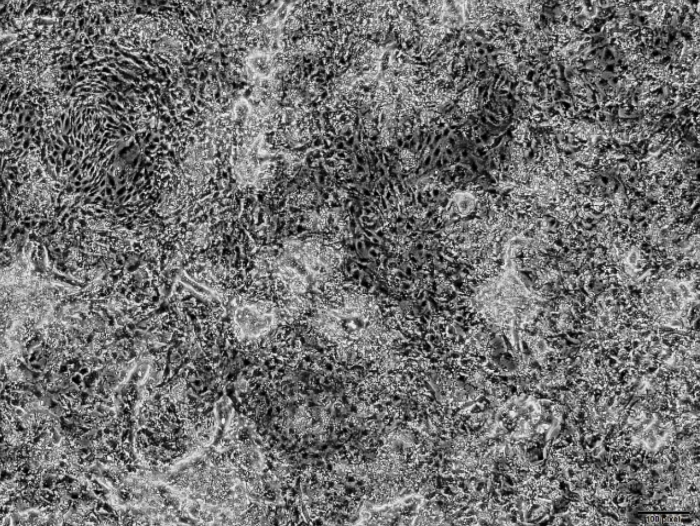
Video 2: Representative video showing an example of a low efficiency differentiation. Please click here to download this video.
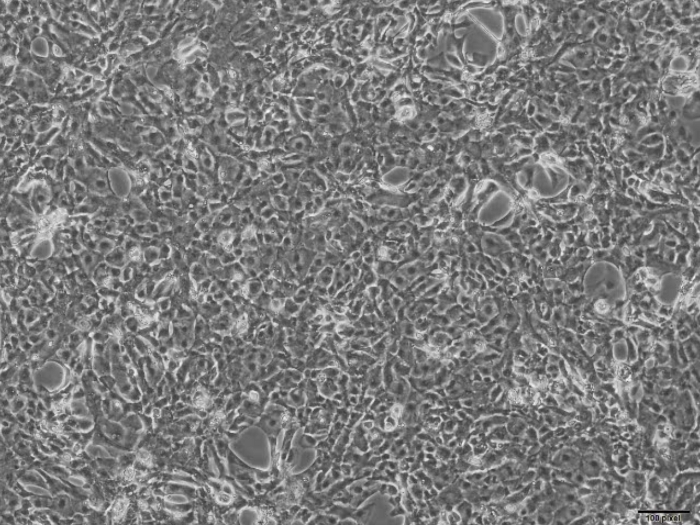
Video 3: Representative video showing an example of a homogeneous monolayer distribution of iPSC-CMs on a glass coverslip. This video shows the recommended cell density to be used for functional analysis. Please click here to download this video.
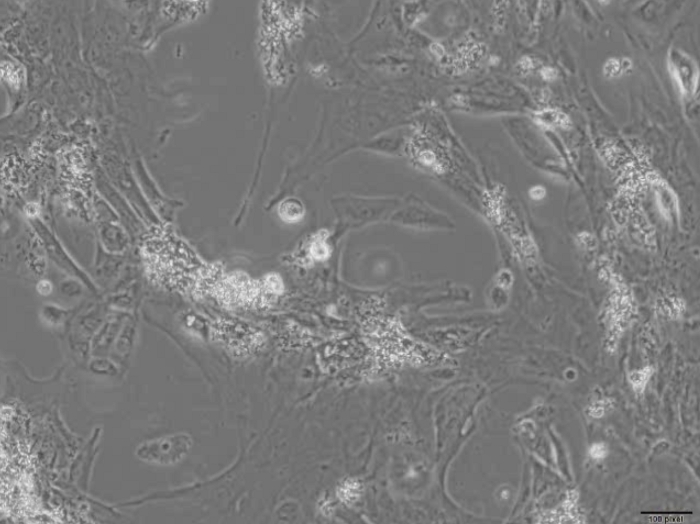
Video 4: Representative video showing an example of cells from a low efficiency differentiation plated onto a glass coverslip. The cells in the video were obtained from a low efficiency differentiation. Cells from such differentiations are usually not distributed homogeneously across the coverslip and it is difficult to consistently find beating cells. Please click here to download this video.
